

The cross-section of a barley root
Ranunculus Root Cross Section
Large, mature tree roots above the soil
Wikimedia Commons has media related to .
Wikiquote has quotations related to Root.
- Botany – University of Arkansas at Little Rock
- Time-lapse photography of root growth on YouTube
This article is about the part of a plant. For other uses, see Root (disambiguation).
«Rooted» redirects here. For the 1969 play and TV movie adaptation, see Rooted (film). For the song, see Ciara discography.

Primary and secondary roots in a cotton plant
A true root system consists of a primary root and secondary roots (or lateral roots).
- Root Hair Overview
- An overview of Root Hair formation
- Use in plant breeding
- Acknowledgements
- Smoke molecules cause root hairs to grow
- Discussion
- Is the lack of root hairs compensated by an increased investment in root growth in general and more specifically in the growth of fine roots?
- Is the role of root hairs for anchorage causing an adaptation in root system architecture?: how does this relate to soil exploration?
- Is the difference between wild-type and mutant larger in a substrate with a high sorption capacity, i.e. low mobility of the limiting nutrients?
- Why do we see a high plasticity with respect to substrate, but only small compensation for the lack of root hairs?
- System limitations — relevance for field conditions
- Discussion
- Root system architecture (RSA)
- Terms and components
- Rights and permissions
- Abbreviations
- Ethics declarations
- Conflict of interest
- Data availability
- Use in plant propagation
- Shade avoidance response
- Genetically transformed cultures
- Additional information
- Publisher’s note
- About this article
- Cite this article
- Keywords
- Root hairs increase the root surface area
- Results
- Shoot and root growth, P acquisition
- Root system architecture in 4D
- Root length density (RLD)
- Root diameter distribution: mean root diameter
- Root distance maps
- Rhizosphere volume fractions (RVF)
- Results
- Model validation
- Root-soil water dynamics under variable precipitation conditions
- Root solute uptake from uniform P distribution with depth
- Root nutrient uptake from declining P distribution along the depth
- Impact of water dynamics vs root hairs on total P uptake
- Impact of root hairs over several years
- Conclusion
- Theoretical considerations
- Overview of the significance to root hairs in P acquisition
- Conclusions
- Author information
- Authors and Affiliations
- Contributions
- Corresponding author
- Funding
Stilt roots of Maize plant
Cross section of an adventitous crown root of pearl millet (Pennisetum glaucum)
Roots forming above ground on a cutting of an Odontonema («Firespike»)
The growing tip of a fine root
Growth from apical meristems is known as primary growth, which encompasses all elongation.
Secondary growth encompasses all growth in diameter, a major component of woody plant tissues and many nonwoody plants. For example, storage roots of sweet potato have secondary growth but are not woody. Secondary growth occurs at the lateral meristems, namely the vascular cambium and cork cambium. The former forms secondary xylem and secondary phloem, while the latter forms the periderm.
Tree roots at Port Jackson
Fluorescent imaging of an emerging lateral root.
Root hair, or absorbent hairs, are outgrowths of epidermal cells, specialized cells at the tip of a plant root. They are lateral extensions of a single cell and are only rarely branched. They are found in the region of maturation, of the root. Root hair cells improve plant water absorption by increasing root surface area to volume ratio which allows the root hair cell to take in more water. The large vacuole inside root hair cells makes this intake much more efficient. Root hairs are also important for nutrient uptake as they are main interface between plants and mycorrhizal fungi.
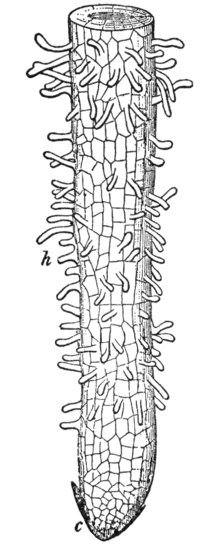
Root tip, showing young root hairs
Root Hair Overview
Root hairs are tiny, hair-like structures that grow on the surface of the plant roots. Their main function is to increase the area available for water absorption and that of minerals and other nutrients. Root hairs are delicate structures that can only survive for a couple of days. They do not have the ability to turn into roots.
Root hairs are attached at the root epidermal cells, and in addition to allowing an affective increase of surface area for water and nutrient uptake they also enlarge the volume of the availalble exploited soil, and aid in the anchoring the plant within the soil.
The formation of root hairs takes place as a series of developmental processes that is initiated with cell fate specification in the meristems.
An overview of Root Hair formation
The Trichoblast root hair forming epidermal cells participate with the diffuse growth phase that is allied to the main root axis elongation.
Growth is reorganized when the fully elongated trichoblast have exited the elongation zone and become localised to the side upon the root hair initiation process.
The plant cells of root hairs can be distinguished cytologically by their delayed vacuolation or dense cytoplasm in early development, near the onset of elongation. Visibility of the root hairs occurs during the swelling at the apical end of the elongated epidermal cells.
In summary the general functions provided by root hairs, that is to say the collection of huge amounts of nutrients and water in the soil can greatly help with plant maintenance and growth.
Cross section of a mango tree
The distribution of vascular plant roots within soil depends on plant form, the spatial and temporal availability of water and nutrients, and the physical properties of the soil. The deepest roots are generally found in deserts and temperate coniferous forests; the shallowest in tundra, boreal forest and temperate grasslands. The deepest observed living root, at least 60 metres below the ground surface, was observed during the excavation of an open-pit mine in Arizona, USA. Some roots can grow as deep as the tree is high. The majority of roots on most plants are however found relatively close to the surface where nutrient availability and aeration are more favourable for growth. Rooting depth may be physically restricted by rock or compacted soil close below the surface, or by anaerobic soil conditions.
. A research group led by scientists at the Technical University of Munich found that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be useful to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM")
Seedlings of thale cress (Arabidopsis). A research group led by scientists at the Technical University of Munich found that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be useful to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM
The roots of plants can do a lot of things: They grow in length to reach water, they can bend to circumvent stones, and they form fine root hairs enabling them to absorb more nutrients from the soil. A team of researchers led by scientists at the Technical University of Munich (TUM) has now identified an important regulator of this process.
A team of researchers led by Caroline Gutjahr, professor for plant genetics at the TUM School of Life Sciences Weihenstephan, has now discovered that it also plays an important role in regulating the growth of roots.
- Baldocchi DD, Xu L (October 2007). «What limits evaporation from Mediterranean oak woodlands–The supply of moisture in the soil, physiological control by plants or the demand by the atmosphere?». Advances in Water Resources. 30 (10): 2113–22. Bibcode:2007AdWR…30.2113B. doi:10.1016/j.advwatres.2006.06.013.
- Brundrett, M. C. (2002). «Coevolution of roots and mycorrhizas of land plants». New Phytologist. 154 (2): 275–304. doi:. PMID 33873429.
- Clark L (2004). «Primary Root Structure and Development – lecture notes» . Archived from the original on 3 January 2006.
- Coutts MP (1987). «Developmental processes in tree root systems». Canadian Journal of Forest Research. 17 (8): 761–767. doi:10.1139/x87-122.
- Raven JA, Edwards D (2001). «Roots: evolutionary origins and biogeochemical significance». Journal of Experimental Botany. 52 (Suppl 1): 381–401. doi:10.1093/jxb/52.suppl_1.381. PMID 11326045.
- Schenk HJ, Jackson RB (2002). «The global biogeography of roots». Ecological Monographs. 72 (3): 311–328. doi:10.2307/3100092. JSTOR 3100092.
- Sutton RF, Tinus RW (1983). «Root and root system terminology». Forest Science Monograph. 24: 137.
- Phillips WS (1963). «Depth of roots in soil». Ecology. 44 (2): 424. doi:10.2307/1932198. JSTOR 1932198.
- Caldwell MM, Dawson TE, Richards JH (1998). «Hydraulic lift: consequences of water efflux from the roots of plants». Oecologia. 113 (2): 151–161. Bibcode:1998Oecol.113..151C. doi:10.1007/s004420050363. PMID 28308192. S2CID 24181646.
Use in plant breeding
The findings not only promote a more thorough understanding of how plants work, but can also be important for future sustainable agriculture.
«With this new and better understanding, of which molecular mechanisms play a role in the development of roots and root hairs, we can breed plants which are better able to absorb nutrients and water from the soil,» Caroline Gutjahr says. «This can for example help crops to resist longer periods of drought, which may occur more frequently in the future.»
More information:
José Antonio Villaécija-Aguilar et al, SMAX1/SMXL2 regulate root and root hair development downstream of KAI2-mediated signalling in Arabidopsis, PLOS Genetics (2019). DOI: 10.1371/journal.pgen.1008327
Journal information:
PLoS Genetics
Acknowledgements
This project was carried out in the framework of the priority programme 2089
Smoke molecules cause root hairs to grow
To check their hypothesis, the team of researchers exposed young arabidopsis plants to the karrikin molecules as they occur in smoke. This indeed significantly increased the growth of root hairs.
«Our results are also interesting in the light of evolution,» Caroline Gutjahr says. «It is likely that KAI2 was initially responsible for controlling certain development processes in all plants, such as, for example, the development of root hairs and similar structures, in reaction to a plant hormone that is so far unknown. In the course of evolution of fire-activated seeds, KAI2 thas presumably developed the additional feature of detecting smoke.»
. A research team led by Prof. Dr. Caroline Gutjahr, Professorship of Plant Genetics at the Technical University of Munich, has discovered, that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be used to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM")
Seedlings of thale cress (Arabidopsis). A research team led by Prof. Dr. Caroline Gutjahr, Professorship of Plant Genetics at the Technical University of Munich, has discovered, that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be used to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM
Discussion
For the discussion part of our work, we will attempt to answer our original hypotheses stated in the introduction.
Is the lack of root hairs compensated by an increased investment in root growth in general and more specifically in the growth of fine roots?
Despite employing two complementary root system architecture measurements (µCT, destructive sampling), we did not observe an increase in fine root growth as a compensation for the lack of roots hairs. This is surprising, as the mutant exhibited a plastic response in root growth with respect to substrate. This finding is in contrast to the one by Klamer et al. (2019) who compared Zea mays WT to the root hair defective mutant rth2. They reported a shift towards finer roots for the mutant. However, in order to detect this shift, they had to compile data across treatments differing vastly in P and water supply. Their plants were growing in subsoil material and were non-mycorrhizal, while the plants in the present experiment showed first signs of mycorrhizal colonization (Table S1) despite the early growth stage. A more intense infection with mycorrhizal fungi as a compensation for the lack of hairs was suggested by Li et al. (2014) for barley and confirmed by Kumar et al. (2019) for maize, but only for later growth stages. The type of mutation is another potential explanation for the differences in compensation mechanism observed by Klamer et al. (2019) and the present experiment. In mutant rth3 the mutated gene encodes a GPI-anchored COBRA like cell wall protein involved in the organization of the synthesized cellulose (Hochholdinger et al. 2008). For the rth2 gene the mechanism is not yet identified.
In general, a significantly higher investment in root growth by the root hair defective mutant as it is reported in literature (Dodd and Diatloff 2016; Klamer et al. 2019) is also found in the present study, although only in relative but not in absolute terms and only for loam, i.e. the substrate with lower P mobility. The shift in root:shoot ratio was not sufficient to compensate for the lack of hairs as total P uptake was significantly lower for rth3 as compared to WT in loam. With respect to physiological plasticity our results are inconsistent. While without morphological and physiological compensation lower uptake rates per unit root are expected, we found no significant differences in normalized P uptake between the genotypes, with only a tendency towards lower values for rth3 in the substrate with the lower P mobility. In general, much lower P uptake rates per unit root surface were found for loam as compared to sand, despite the low plant P status in loam (leaf tissue P concentration of 2.4 mg g−1 for rth3 and 2.6 mg g−1 for WT, which is expected to trigger expression of high affinity P transporters. This is in line with relative root gene expression data for the same experimental setup, reported by Ganther et al. (2021). They did not find gene functions relating to phosphate uptake to be differentially expressed between WT and rth3. It is still possible that the activity of the transporters is regulated at the protein level, since apart from transcriptional regulation, post-translational modifications of phosphate transporters are widespread (Vetterlein and Tarkka 2018). For soil based systems, uptake rate per unit root is strongly influenced by diffusion and root to soil contact. Haling et al. (2013) found significantly lower P uptake rates per unit root length for the root hair defective mutant of barley compared to the wild-type, for high but more so for low P soils. Root hairs were relevant for maintaining contact in loose soils and for improved penetration in dense soils. The importance of hairs for P uptake in particular under conditions of restricted P transport is confirmed by observations in hydroponics, i.e. systems with no major limitations for transport. Suzuki et al. (2003) found in hydroponic systems no differences in P uptake rates per unit root dry weight between rice genotypes differing in root hair development, irrespective of P supply. The differences between substrates observed here are related to their differences in P diffusion.
Is the role of root hairs for anchorage causing an adaptation in root system architecture?: how does this relate to soil exploration?
Bengough et al. (2016) have demonstrated the importance of root hairs for anchorage using the same maize genotypes as in the present study. Peak anchorage forces were up to five times greater for the wild-type compared to the root hair defective mutant. As a consequence, wild-type primary roots penetrated deeper into the soil during a given time interval as compared to the mutant. The difference was fivefold at low bulk density and decreased continuously with increasing bulk density/mechanical impedance. At the bulk density and water content investigated in the present experiment, penetration resistance in both substrates was well below 0.1 MPa (U. Rosskopf, S. Peth, D. Uteau personal communication). This value is an order of magnitude below the critical value for root elongation rate of 2 MPa suggested by Bengough et al. (2011). With two exceptions no differences were observed in depth exploration between the two genotypes within the same substrate. At 7 DAP, wild-type showed significantly higher values in the lower depth intervals, which is in line with the postulated role of hairs for anchorage, however in absolute values these differences were very small. A distinct exception occurred for the last time point in loam most likely due to a technical artefact. During the last four days of growth, plant water consumption for wild-type in loam was so high, that short-term desiccation of top soil occurred between the watering events. This likely caused root shrinkage which in turn reduced recovery with X-ray CT. The study of Bengough et al. (2016) focused on the very early growth stage, i.e. 3 days after germination with just the primary root (1–3 cm long) at the start of the experiment and a duration of the experiment of maximum 48 h. At later growth stages, it can be expected that lateral root formation as well as seminal roots partly take over the function of anchorage from the hairs (Bailey et al. 2002). Lateral roots on the primary root are abundant at 7 days after planting (Fig. S5).
Root distance histograms were derived from X-ray CT data to quantify soil exploration in more detail, as they simultaneously take into account the actual 3D geometry, differences in length and diameter (Schlüter et al. 2018). Root distance histograms or the mean root distance derived from these data (Figs. 7, 8) are a very sensitive measure in particular at early growth when exploration is poor. This is indicated by the strong impact of delayed lateral root formation in two out of six replicates being reflected in mean root distance but not in RLD (Fig. S5). While the measure nicely reflects the progressive exploration of the soil columns from top to bottom and over time with distinct differences between substrates, no significant differences between genotypes were observed. At later growth stages, when root length densities were higher a limit in mean root distance of 3 mm was attained. Such a limit in the range of 3 mm was also reported by Lucas et al. (2019) for root-induced biopores in undisturbed field samples. This suggests that an investment in more root growth may not pay off in terms of better root exploration but only increase competition between roots for the same resources. However, whether that is really the case would be better assessed with nutrient specific rhizosphere volume fractions, which are more sensitive to growth patterns at high RLD.
For the RVF, for which the measured root hair length (0.24 mm) was explicitly added to the assumed extension of the P depletion zone (1.8 mm derived from Hendriks et al. (1981)) for the wild-type, no significant effect of genotype was detected at 14 and 21 days after planting at all depths. RVF reached 25 to 50% at day 21 (Fig. 9), indicating that a large fraction was already explored for P at this early stage. Note that these results are hypothetical and depend on the assumed extension of rhizosphere. More accurate RVF estimates would require spatially resolved information about radial depletion patterns of plant available P on multiple, intact rhizosphere sections to capture them representatively.
It should be noted that the measured root hair length of 0.24 mm is rather short as compared to literature data. Frequently higher values in the range of 0.7 to 0.9 mm are reported for maize (Hendriks et al. 1981; Weber et al. 2018). In general root hair lengths can vary with soil P status and bulk density (Haling et al. 2013; Jungk 2001); there was a tendency towards longer root hairs in sandy substrate. Increasing hair length in our calculation of RVF would result in higher values (Fig. S6). This increase is linear for realistic root lengths. The relative importance of root diameter over root hair length for RVF would increase as the extension assumed for the rhizosphere process in question decreases.
Root length density is the dominant factor that governs differences in RVF. However, genotype (i.e. added hair length) and substrate had an additional impact (Fig. S7). For the latter one cannot disentangle the preferential growth along the wall in loam (fewer neighbouring soil voxels for roots at the wall) from the differences in root diameter (increasing the number of neighbouring soil voxels with circumference) between loam and sand. These results are valid irrespective of the exact value for the hypothetical spatial extent of P depletion, i.e. for the only variable that could not be measured in our study.
Is the difference between wild-type and mutant larger in a substrate with a high sorption capacity, i.e. low mobility of the limiting nutrients?
Nutrient mobility is expected to be lower in loam as compared to sand as loam has a higher number of sorption sites based on higher content of Fe-oxides, clay and organic matter (Vetterlein et al. 2021). Interpretation of biomass nutrient concentration per se is confounded by dilution through growth. Interpretation of shoot nutrient uptake is confounded by differences in shoot size and hence nutrient requirement. To evaluate differences in mobility we therefore used not only P uptake itself, but in addition the stoichiometric ratio of Ca (having a high mobility in soils) over P (having a low mobility in soils) (Ågren and Weih 2012). For our substrates Ca:P ratio was well suited to show the differences between genotypes for loam. Differences between genotypes were significant for most of the measured growth and uptake parameters in loam, but not in sand. This is in line with our hypothesis and the observation of others (Haling et al. 2013; Suzuki et al. 2003), i.e. roots hairs only matter if transport to the root surface is limiting uptake.
Why do we see a high plasticity with respect to substrate, but only small compensation for the lack of root hairs?
The most frequent cause for shifts in root diameter reported in literature is alterations in soil compaction, bulk density and mechanical impedance, which are tightly linked with changes in soil water content and gas diffusion (Bengough et al. 2006, 2011; Clark et al. 2003; Colombi and Walter 2016; Correa et al. 2019). Root diameter increase by up to twofold in case of mechanical impedance has been reported, as a result of cortical cells expanding radially due to microfibril reorientation in the primary cell wall (Bengough et al. 2011). Causal relationship is straight forward, if increasing bulk densities within the same soil/substrate are investigated (Haling et al. 2013; Tracy et al. 2012). An increase in root diameter upon compaction was also observed for our loam in a parallel experiment (Table S2). When comparing different substrates causal relationships are more difficult to unravel. Kirby and Bengough (2002) have nicely shown for the comparison of a sandy loam and a clay loam that penetration resistance alone is not sufficient to predict root thickening. They demonstrated that local values of axial and shear stresses experienced by the root near its tip may be as important as penetration resistance in constraining root growth. Ethylene is often associated with the morphological response of roots to mechanical impedance (Clark et al. 2003; Dreyer and Edelmann 2018). Increased levels of ethylene have been observed to induce increase in root diameter even for unimpeded roots (Baluška et al. 1993). An induced expression of genes related to phytohormone signalling was detected only in sand by Ganther et al. (2021) in an experimental setup like the one used here. Ethylene, but also gibberellic acid and jasmonate were affected, which indicates that processes related to development and growth are altered by the substrate. Further studies are required to evaluate whether observed differences can be explained by mechanical properties beyond mechanical impedance (Kirby and Bengough 2002), root (Ma et al. 2013; Pandey et al. 2021) or microbiome ethylene production (Zhang et al. 2020) or differences in diffusion processes within the root or in the rhizosphere (Hartman 2020). Anoxia as a trigger for ethylene production is ruled out for our system, at least not beyond occasional microsites. At the volumetric water content used in this experiment (22% in loam, 18% in sand), air filled pore volume is well above 10% even at the bottom of the sand columns (Vetterlein et al. 2021).
Increase in root diameter in sand as compared to loam did not result in an increased investment in root growth in general. Root:shoot ratio was lower in sand. This suggests that plant demands in terms of water and nutrients could be covered with a less intensive soil exploration. As differences in P uptake did not occur in sand it is difficult to assess whether compensation for the lack of root hairs did not occur in sand because there was no need for it or because fine root growth was hindered by other factors in this substrate. Unfortunately, no support for either possibility can be derived from the data of Klamer et al. (2019) as they have evaluated changes in diameter across all treatments including two textures.
System limitations — relevance for field conditions
Discussion
At the field scale, the modelling in this paper has demonstrated a large impact of the presence of root hairs and environmental conditions on P uptake by plants. We enhanced a model that considers root-soil interaction at the field scale by considering unsaturated soil moisture conditions and solute transport.
A qualitative model evaluation illustrated the validity of our modelling methodology (Fig. 4). Saturation trends where both spatially and temporally similar, which suggests that our root water uptake model was sufficiently accurate for predicting field scale dynamics under partially saturated conditions (Fig. 4 (a)). Similarly, our model was also able to capture the influence that the roots have on soil solution P concentrations (Fig. 4 (b)). While these estimates were more qualitative than the saturation comparison, the influence that roots have on taking up dissolved P impact the P distribution in the soil in a similar manner. The model did not predict the exact saturation magnitudes due to differences in soil physical properties, root distribution, and unknown lower boundary conditions. However, if needed, our model could be fit directly to the specific data, possibly allowing us for inverse modelling to estimate the soil physical properties and root suction pressures given information about the root distributions. Our model could likely be fit to the exact P profiles measured in these studies as well. However, this was outside of the scope of our study. The model validation grants us more confidence in our models ability to predict water and P uptake from field systems.
For the basic scenarios, the model was able to deal with early and late precipitation scenarios (Fig. 5). The dynamic differences considering the early (Fig. 5 (a)) vs late (Fig. 5 (b)) scenarios illustrate some differences in the dynamics of the soil moisture along the profile. Both profiles appear characteristically the same after 90 days. However, while the profiles are qualitatively the same, the water uptake from the two different scenarios differs drastically (Fig. 6). Though soil water content with depth under different rainfall scenarios may appear similar after 90 days, the preceding dynamics are very different. Information regarding the dynamics of soil water movement cannot be extracted from the steady state soil water content profile measured at 120 days, but must consider changes over time.
Although P concentrations change marginally over a single growing season, simulation that were run for several growth cycles under realistic precipitation patterns revealed a noticeable decrease in soil P concentrations for soils containing plants with root hairs (Fig. 11). Most UK soils have abundant P reserves, but fertiliser is still applied as most P is adsorbed to soil and unavailable to plants. Root hairs clearly enhance the uptake of this P, leading to potentially greater nutrient use efficiency and decreased needs for fertiliser application. Taking barley as an example, which is grown on 1× 106 ha of land in the UK, the 0.001 molp m−3 difference in P that we found between plants with and without root hairs (after six growing seasons), equates to over 150 t of P captured from the soil. The same area of land has 24,000-24,500 t of P added to it each year (DEFRA 2017), which amounts to 144,000-147,000 t of P over the 6 growing seasons. The total quantity of P taken up by the root hairs amounts to less than 0.1% of the P input. Thus, while root hairs enhance plant P acquisition, the effect of root hairs poses no risk of mining soils under current production practices.
Despite the limited changes of P concentrations down the soil profiles, the P uptake rates by the roots were significantly different under the precipitation conditions and the initial quantities of P in the soil (Fig. 9). The influence of the root hairs was modelled to account for 50% of the total P taken up by the plant root. This was in accordance with previous experimental literature (Brown et al. 2000; Keyes et al. 2013). These results appear consistent with modelled P uptake trends that only considered solute transport in soil (Itoh and Barber 1983). Image based modelling of root hair enhanced P-uptake also estimated that root hairs account for up to 50% the total P uptake (Daly et al. 2016). Our field scale model results are similar to these image based results. The P uptake rate by the roots scaled by 70% as a result of soil P concentrations reduction by 70% (comparing Fig. 9 (a-b) to (c-d)). This is expected, as the root uptake rates (Eqs. (26), (27), and (30)) are related to the P concentration in the soil via Michaelis-Menten kinetics (Nye and Tinker 1977). Furthermore, this is consistent with previous experiments (Brown et al. 2012b). In glasshouse experiments that considered a soil with an initial inorganic P content of 590 mgP kgbulk−1, the amount of P accumulated in the plant shoots nearly doubled after 500 mgP kgbulk−1 was added to the soil (Brown et al. 2012b).
The simulations also revealed the sensitivity of the uptake rates to the precipitation events (Fig. 9 (a, c) vs (b, d)). Phosphorus uptake rates were suppressed by 25-60% if the onset of precipitation was delayed by 60 days. The results from this study appear more pronounced than those found in the image based modelling study (Daly et al. 2016), which only saw marginal changes in the nutrient uptake due to drier conditions. As our field scale model considers large spatial averages, we think that it is more likely that full roots (and root hairs) may be under complete drought conditions, while other roots may still have to access smaller wet subdomains, and these would spatially manifest in greater reductions in the overall root uptake. We also note that the image based modelling study (Daly et al. 2016) maintained fixed soil moisture for their different scenarios, which facilitated nutrient fluxes to the root hairs whilst in our study the soil water saturation was considered dynamic.
Similar experiments considering maize roots have seen similar reductions in the amount of P taken up by plants under variable drought conditions (Resnik 1970); consistent with our model results. The P-uptake theoretically peaks within the first 2 months under wet conditions (Fig. 9), illustrating that the soil water regime plays a considerable role in P use efficiency. While plant yields will likely depend on the quantity of plant available nutrients in the soil, our results are consistent with the claim that plant P use efficiency (in our case, the P uptake rates) is influenced by frequency of precipitation (Silber et al. 2003). Moist conditions during early stages of a growing season would increase plant P use efficiency.
There were various assumptions in the derivation of the model, which focused on the impact of root hairs on P capture and uptake. Overland flow was omitted from this study. Prolonged flooding is not modelled as barley can only survive several days of waterlogging. Future studies could investigate flooding by incorporating the potential-form of Richards’ equation as presented in Duncan et al. (2018), where an approach how to swap between flooded and non-flooded boundary conditions is described. Although evaporation influences the net water fluxes in the soil, we do not explicitly include this process, as the focus of the study is on root soil interactions. To include our model in a broader scale, it would be important to consider different climatic variables (i.e. temperature, relative humidity, and wind speed) in order to estimate surface evaporation as done in previous studies (Heppell et al. 2014). For our study site, where rainfall exceeds evaporation and there is a net eluviation of nutrients to deeper depths, this assumption is fair, but for drier regions it would need to be included.
While the model simplifies the effect that root hairs have under natural environmental conditions, it provides predictions that agree with experimental observations. For example, experimental results suggest that root hairs nearly alleviate the impacts of drought entirely (Brown et al. 2012b). While our model suggests a strong mitigation to the effects of drought (e.g. maintaining significantly higher P uptake rates with root hairs compared to no hair), drought still has a comparative effect on the uptake rates (Fig. 9 (a, c) vs (b, d)). The enhanced resistance to drought may be due to the modification that root hairs impart on the soil structure local to the root interface (Koebernick et al. 2017), thus augmenting the moisture dynamics and, therefore, nutrient fluxes in the rhizosphere. Future work could develop more detailed models that better account for local rhizosphere structures (Daly et al. 2017) in the context of nutrient fluxes and considering how root hairs impact water uptake (Carminati et al. 2017). Similar to how rhizosphere features have impacts on soil chemistry, local microbiota will also be affected by root exudates, which will likely create feedbacks and possible mutualistic benefits (Dupuy and Silk 2016; Kuzyakov and Blagodatskaya 2015). Future work could try to consider the ensemble of these various rhizosphere impacts on the pore scale and how they upscale to the field.
Root system architecture (RSA)
Terms and components
Extrinsic factors affecting root architecture include gravity, light exposure, water and oxygen, as well as the availability or lack of nitrogen, phosphorus, sulphur, aluminium and sodium chloride. The main hormones (intrinsic stimuli) and respective pathways responsible for root architecture development include:
Rights and permissions
Reprints and Permissions
Abbreviations
- CT:
- DAP:
Days after planting
- L:
- RDH:
Root distance histogram
- RLD:
Root length density
rth3
:Root hair defective mutant
- RVF:
Root volume fraction
- S:
- WT:
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Use in plant propagation
This section needs expansion. You can help by adding to it.
Shade avoidance response
Genetically transformed cultures
Additional information
Responsible Editor: Tim S. George.
Publisher’s note
About this article
Cite this article
Lippold, E., Phalempin, M., Schlüter, S. et al. Does the lack of root hairs alter root system architecture of Zea mays?.
Plant Soil 467, 267–286 (2021). https://doi.org/10.1007/s11104-021-05084-8
Received
Accepted
Issue Date
Keywords
Root hairs increase the root surface area
To achieve a large surface area through which water and nutrients can be absorbed, the roots of plants grow fine root hairs. José Antonio Villaécija-Aguilar, a Ph. D. student in Caroline Gutjahr’s team, has now made the discovery that KAI2 is both necessary for the growth of this root hair and for the downward growth of roots.
«It is likely that this not only applies to the thale cress (Arabidopsis), which can be found almost everywhere in the world and is used by us as a model plant,» Caroline Gutjahr says, «but possibly also for many other plants, for example cereal crops.»
. A research team led by scientists at the Technical University of Munich has discovered, that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be used to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM")
Seedlings of thale cress (Arabidopsis). A research team led by scientists at the Technical University of Munich has discovered, that the KAI2 protein regulates essential functions of root and root hair growth. The findings not only promote a more thorough understanding of how plants work, but can also be used to breed plants which are better able to absorb nutrients and water from the soil. Credit: Andreas Battenberg/TUM
Results
Shoot and root growth, P acquisition
Plant P tissue concentration was low in both substrates as intended in order for root hairs to play a role in P acquisition under P limited conditions (in loam 2.4 mg g−1 for rth3 and 2.6 mg g−1 for WT, in sand 2.7 mg g−1 for rth3 and WT). Overall, there was a significant impact of substrate on shoot and root growth, with a shift in root:shoot ratio (Fig. 2a, b, c). Lack of root hairs resulted in a reduction of shoot and root growth. These effects were larger for shoots than for roots, the latter being reflected in a shift in root:shoot ratio towards the roots for rth3 (Fig. 2c). Growth reduction (shoot and root) was larger for loam than for sand and the differences between genotypes were even more obvious for plant P content (Fig. 2d). There was no significant difference between the genotypes with respect to P uptake per unit root surface, albeit there was a tendency for lower uptake for rth3 as compared to WT for loam (Fig. 2e). Likewise the Ca:P ratio showed higher values for rth3 as compared to WT for loam. However, no difference between genotypes was found for sand (Fig. 2f).
A higher investment in root growth to compensate for the lack of absorbing surface provided by root hairs was not found in absolute terms (Fig. 2b) but in relative terms, at least for loam (Fig. 2c).

Impact of substrate (loam, sand) and maize genotype (wild-type—WT, root hair defective mutant rth3–rth3) on shoot dry weight (a), root length (b), root:shoot ratio (c), shoot P content (d), P uptake per unit root surface (e) and the stoichiometric ratio of the mobile element Ca over the immobile nutrient P (f) in the shoot 22 days after planting
Full size image
The X-ray dose associated with X-ray CT scanning had no significant impact on shoot or root growth with the scan settings and scanning frequency chosen (Fig. S1).
Root system architecture in 4D
Time resolved X-ray CT scanning and superposition of scans from different time points provided insight into the 3D architecture of the root system and its temporal development including detailed information on root diameters (Fig. 3). Comparison with the sketch of maize root development clearly shows that, with X-ray CT, the primary roots and seminal roots including their laterals can be identified (pink colour—7 days). The roots captured additionally at day 14 (green) mainly represent the crown roots and their laterals, while those captured at day 21 represent the brace roots (blue). It should be noted that this simple assignment between scanning events and root type is only valid for the main root axis and differs in detail for the lateral roots. Note that the recovery of roots with X-ray CT was different for the loam and sand and that this difference should be kept in mind for the interpretation of the X-ray CT acquired results. The recovery of roots was equal to 99% (R2 = 0.84; n = 24) for the sand treatment whereas it amounted to 71% (R2 = 0.61; n = 19) for the loam treatment (Bulk density g cm−3).

Root system architecture (from − 1.27 to 14.77 cm depth) derived from X-ray CT scanning at 7, 14 and 21 days after planting (pink = 7 DAP; green = roots grown between 7 and 14 DAP; blue = root grown between 14 and 21 DAP). A representative example for WT (root length of the sample closest to the mean of the six biological replicates per treatment) is shown in both substrates. The sketch in black and grey illustrates the different root types of maize which can be found
Full size image
Root length density (RLD)
Root length density profiles (Fig. 4, Fig. S3) showed significant differences between substrates for most depth intervals at 14 DAP and the lower ones at 21 DAP. Genotype only had a significant impact on depth exploration at 7 DAP, when overall RLD was still very low. For 21 DAP significant impact of genotype in the upper depth intervals is related to desiccation induced artefact described below. Hence, only the differences found for the lower depth intervals will be discussed further. The strong increase in root length density in the lower part of the columns observed for loam towards the end of the experiment was mostly outside the scanned region. This was quantified by destructive sampling and analysis with WinRhizo at harvest (Fig. S3).

Change of root length density with depth for 7, 14 and 21 days after planting for two maize genotypes (wild-type—WT, root hair defective mutant rth3–rth3) grown in loam (L) and sand (S). Data are derived from X-ray CT scanning; n = 6, shaded areas represent standard error. Statistics: two-factorial ANOVA in conjunction with Tukey’s HSD test was conducted for depth interval. Significant effect of factor is denoted by s for substrate, g for genotype and x for interaction, for p > 0.05 no letter is displayed
Full size image
It should be noted that, due to lower recovery of roots in loam than in sand, the root length densities for the loam treatments were underestimated relative to the sand treatments for X-ray CT derived data (Figs. S2, S3). This was especially true for L_WT on day 21 with the highest proportion of roots < 100 µm (Fig. S4).
Very likely, a higher root length density at the top of the column for the L_WT treatment at 21 days was not recovered due to the stronger desiccation of the soil, which might have led to root shrinkage. The associated reduction of root diameter could be responsible for a lower recovery of laterals during root segmentation (Fig. 4).

Depth distribution of roots younger than 7 days at 14 days and 21 days after planting for two maize genotypes (wild-type—WT, root hair defective mutant rth3–rth3) grown in loam (L) and sand (S). Data are derived by simple arithmetic operations on the dataset shown in Fig. 4; n = 6, shaded areas represent standard error. Statistics: two-factorial ANOVA in conjunction with Tukey’s HSD test was conducted for depth interval. Significant effect of factor is denoted by s for substrate, g for genotype and x for interaction, for p > 0.05 no letter is displayed
Full size image
Root diameter distribution: mean root diameter

Change of mean root diameter with depth for 7, 14 and 21 days after planting for two maize genotypes (wild-type—WT, root hair defective mutant rth3–rth3) grown in loam (L) and sand (S). Data are derived from X-ray CT scanning; n = 6, shaded areas represent standard error. Statistics: two-factorial ANOVA in conjunction with Tukey’s HSD test was conducted for depth interval. Significant effect of factor is denoted by s for substrate, g for genotype and x for interaction, for p > 0.05 no letter is displayed
Full size image
Root distance maps
The exploration of the soil by roots can be visualized with root distance maps (Fig. 7a) and quantified with root distance histograms (RDH) (Fig. 7b). The mean root distance in soil is derived from that RDH and differentiated according to soil depth (Fig. 8). In general, the mean root distance reflects the root length density rather well (Fig. 4). That is, an increase in root length density results in a higher frequency of short root distances making large distances less frequent (Fig. 7b) and hence reducing mean root distance (Fig. 8). Seven days after planting the root network is poorly developed at the bottom of the field of view and comprises only the primary root and a few seminal roots without laterals, which causes a marked increase of mean root distance with depth. At this early stage the same root length densities in sand and loam evoke different mean root distances across the entire column. This is due to two out of six replicates which had no laterals yet along the primary root at this time point for treatment S_WT (Fig. S5). Their absence has a huge impact on mean root distance in a sparsely populated soil (7 DAP) that is not reflected to the same degree in RLD. In addition, sand and loam treatments might differ in the spatial arrangement of seminal roots. The seminal roots and the primary root seemed to be more clustered in one semicircle of the column wall in loam as compared to more equidistant radial positions in sand (Fig. S5). At 14 DAP the root length density is higher in sand for almost the entire field of view except for the very bottom (below − 12 cm). This difference in RLD was also reflected in the corresponding depth distribution of mean root distance, i.e. shorter mean root distance with a higher RLD and vice versa. At 21 DAP the field of view is already densely populated with roots in both substrates. There seems to be a universal limit at approx. 3 mm below which the mean root distances cannot fall despite different RLD in the range of 4–8 cm cm−3. For all scanning events differences in mean root distance between genotypes are absent in both substrates, except for the lowest depth intervals at 7 DAP.

a Root distance maps determined 7, 14 and 21 DAP for the S_WT sample depicted in Fig. 3. b The root distance histograms are shifted towards shorter distance with increasing root length density over time
Full size image

Depth profile of mean root distance for 7, 14 and 21 days after planting for two maize genotypes (wild-type—WT, root hair defective mutant rth3–rth3) grown in loam (L) and sand (S). Data are derived from frequency distribution of distances derived from distance maps calculated on 3D X-ray data; n = 6, shaded areas represent standard error. Statistics: two-factorial ANOVA in conjunction with Tukey’s HSD test was conducted for depth interval. Significant effect of factor is denoted by s for substrate, g for genotype and x for interaction, for p > 0.05 no letter is displayed
Full size image
Rhizosphere volume fractions (RVF)
We recall that the hypothetical rhizosphere volume fractions (Fig. 9) are directly derived from the root distance histograms by determining the frequency of soil voxels with root distances < 1.8 mm, which we considered to be a typical rhizosphere extent for P. In addition, for root hairs of the maize wild-type, the rhizosphere extent was increased by the measured root hair length of 0.24 mm. Again, the vertical distributions of rhizosphere volume fractions reflect root length density profiles for all time points very well. The only deviation from this congruence is a much higher RVF in the top 5 cm at 21 DAP in sand despite similar RLD values in that depth. This increase in RVF was not exclusively due to the larger root diameter in sand, as this would have led to more soil voxels in the direct vicinity of the root interface in the entire scanned region and not just the top. The insets at 21 DAP (Fig. 9) show vastly different RVF (orange) for one loam (a) and one sand (b) column with identical RLD. Roots in loam had a preference for growing along the wall, supposedly in cracks that formed due to desiccation. The rhizosphere of roots growing along the wall was truncated to a semi-circle and contributed less to the RVF. Despite explicitly accounting for hair length, genotype had no significant effect on RVF.

Depth profile of rhizosphere volume fraction for 7, 14 and 21 days after planting for two maize genotypes (wild-type—WT, root hair defective mutant rth3–rth3) grown in loam (L) and sand (S). Data are derived from frequency distribution of distances derived from distance maps calculated on 3D X-ray data; Rhizosphere volume fraction is determined assuming a typical rhizosphere extent of < 1.8 mm for P depletion reported by Hendriks et al. (1981) for 5 day old root segments. For the wild-type 0.24 mm were added to account for hair length; n = 6, shaded areas represent standard error. Statistics: two-factorial ANOVA in conjunction with Tukey’s HSD test was conducted for depth interval. Significant effect of factor is denoted by s for substrate, g for genotype and x for interaction, for p > 0.05 no letter is displayed
Full size image
Results
Model validation

Qualitative model evaluation. a Comparing trends in saturation degree profile trends between the model output and measured field observations (Volaire and Thomas 1995) for two separate time periods (near the beginning of the experiment (days 0-5) and in the middle of the experiment (days 28-30). b Comparing the general trends between modelled and measured normalized P concentrations in solution (Gahoonia et al. 1994)
Full size image
Root-soil water dynamics under variable precipitation conditions
The resulting profiles for the two separate rainfall simulations are illustrated in Fig. 5. Using continuous precipitation initialized from t= 0 days (Fig. 5(a)), the simulations resulted in a gradual increase in the overall soil water content along the soil profile. After 60 days, water content had noticeably decreased near the bottom of the rooting zone. By 90 days, the soil water content near the root tip (˜ 0.5 m) flattened out near the residual water content, as the plant root suction can no longer exceed the soil matric potential. Different characteristic behaviour was observed in Fig. 5(b), where the initialization of the rain begins at t= 60 days and reached steady state at t = 75 days (simulated as a smooth step function where smoothing was made over the 15 day period). The profile initially dries from the soil surface down to the depth just behind the root tip location (0-30 days). On day 60, the soil surface begins to wet due to the precipitation. By days 90 and 120, the profiles are the same for precipitation at t = 0 or t = 60 days.

Full size image

Total root water uptake rate as a function of time. Water uptake was integrated along the rooting depth and around the rooting zone (Eq. (22)) for two precipitation scenarios. The blue curve illustrates the root water uptake dynamics considering the scenario with immediate and constant irrigation from the beginning of the simulation. The red curve illustrates the root water uptake dynamics in response to zero irrigation for the first 60 days, increasing as a smoothed step to reach steady state by day 75 (plotted in the purple highlighted region with the axis on the right)
Full size image
Root solute uptake from uniform P distribution with depth
Profiles of the total P concentrations based on an initial uniform distribution down the soil depth are plotted in Fig. 7. Root P uptake over the course of 150 days demonstrates small changes in the overall P concentrations. The overall P in the domain is less for the simulations influenced by root hairs. Reduction in P profiles appears more pronounced in the cases where rain is initiated at train = 0 days (Fig. 7(a-b)) compared to the delayed rain scenarios (Fig. 7(c-d)). It is worth noting that the overall change in the P profiles are marginal for all simulations, resulting in a maximum percent change below 2% of the initial value.

Full size image
Root nutrient uptake from declining P distribution along the depth
Simulation results that have soil P concentration decreasing exponentially with depth were plotted as percent differences on Fig. 8 in order to provide a clearer comparison of the relative changes at each depth. Considering first the conserved volume simulations, the largest percent changes occurred in the t = 0 days rainfall (Fig. 8(a-b)), with over a 0.5% decrease in P at the maximum root depth due to the influence of root hairs. The remaining P in the hairless simulation was consistently greater in magnitude than their wild type counterpart simulations for both excess and conserved distributions in Fig. 8.

Profile of the percent difference of P with respect to the initial values at each depth considering root growth and P uptake for two precipitation scenarios, four initial decaying P distributions, and considering with (ψH=1) and without (ψH=0) root hairs. The scenario in (a) and (b) illustrates the final P profile after 150 days considering immediate and constant irrigation from the beginning of the simulation, while (c) and (d) consider precipitation initiating at t = 60 subsequent to the simulation. We note that the subdomain in the figures focusses on the rooting zone, not the full domain
Full size image
Impact of water dynamics vs root hairs on total P uptake
The total P uptake rates are compared for all of the simulations in Fig. 9. Comparing first the low P vs high P scenario with precipitation throughout the simulations (Fig. 9(a) and (c)), shows that the uptake rates for all of the simulations was lower in the low P scenario than in the high P scenario, proportional to the volume of P in the system. Because the low P scenario has 70% less total soil P than the high P scenario, the reduction from the P uptake rate from the high P scenario (Fig. 9(c)) to the low P scenario (Fig. 9(a)) appears to also scale back by 70%. Uptake rates were, as expected, consistently greatest for the excess distributions, likely attributed to the scaling effect that the uptake rates have with the concentrations of P in the system.

Root phosphorus uptake rate over time as impacted by each of scenarios. Each of the scenarios considers simulations with root hairs (ψH=1) and without root hairs (ψH = 0) for uniform initial phosphorus distribution (solid curves), mass conserved decaying distribution (dashed), and excess mass decaying distribution (semi dashed). Plots in (a) and (b) consider simulations with initially low phosphate content in the soil, while (c) and (d) consider initially high phosphate content in the soil. Simulations in (a) and (c) consider rainfall initiated from the beginning of the simulation, while (b) and (d) consider precipitation after 60 days
Full size image
For all of the scenarios, P that distributed uniformly had lower uptake rates than if P decreased exponentially with depth. This was likely due to a locally increased concentration of P as the root initially grows down into the soil domain. Simulations of the hairless roots consistently resulted in a 50% reduction in the maximum uptake rate in comparison to the hairy root counterpart simulations.
One of the most striking results was the overall change in dynamics when comparing the simulations with constant precipitation throughout the simulations (Fig. 9(a) and (c)) and the simulations with precipitation initiating after 60 days (Fig. 9(b) and (d)). Where the P uptake in Fig. 9(a) and (c) peaks at 15 days, with delayed rainfall simulations (Fig. 9(b) and (d)) uptake rates never achieve the same maximal values. The reduction in the P uptake rate ranged between 30 and 55% for the different scenarios.
The total quantities of P can also be observed by integrating the uptake rates over time (Fig. 10). Within the first 50 days, the root uptake doubles the quantity of P in the immediate rain scenario (Fig. 10 (a) and (c)) as compared to the delayed rain scenario (Fig. 10 (b) and (d)). After the rain initiates in the train = 60 days scenario (Fig. 10 (b) and (d)), the uptake rate rapidly increases. Thus, the difference between in the quantities between the two rain scenarios range between a reduction of 33-35% the total P taken up by the plants.

Total phosphorus uptake by plant roots over time as impacted by each of scenarios. Each scenario considers simulations with root hairs (ψH=1) and without root hairs (ψH = 0) for uniform initial phosphorus distribution (solid curves), mass conserved decaying distribution (dashed), and excess mass decaying distribution (semi dashed). Plots in (a) and (b) consider simulations with initially low phosphate content in the soil, while (c) and (d) consider initially high phosphate content in the soil. Simulations in (a) and (c) consider rainfall initiated from the beginning of the simulation, while (b) and (d) consider precipitation after 60 days
Full size image
Impact of root hairs over several years

Soil P concentration after multiple growing seasons. (a) illustrates the relative impact that hairless mutants would have on the soil P compared to wild type hairy roots (b). Each curve depicts the end of the year P distribution along the soil depth assuming an initially uniform distribution at the beginning end of 2003 and simulating successive seasons up to the end of 2009. We note that the subdomain in the figures focusses on the rooting zone, not the full domain
Full size image
Conclusion
Adaptations in root system architecture in response to lacking root hairs were investigated with a comprehensive experimental setup that combined nutrient uptake analysis, destructive root sampling and X-ray CT scanning that allows monitoring various root system architecture metrics over time. The CT derived metrics enable quantification of soil exploration and at the same time integrate the effect of various root traits, i.e. root diameter, 3D-distribution, depth distribution, hair length.
Experimental conditions were well suited to confirm the general consensus on root hairs being of particular relevance for uptake of low mobility nutrients such as P, especially in soils with a high sorption capacity. Root hair defective mutants showed low plasticity of root traits related to limited P availability, despite their general ability to express high root plasticity. The function of the root hairs for anchoring did not result in different depth profiles of the root length density. We suggest that, in more developed root systems, as in our experiment, part of the anchoring function can be taken over by lateral roots.
Both maize genotypes showed a marked response to substrates differing in soil texture mainly reflected in mean root diameter. Increase in root diameter is typically induced by higher penetration resistance. However, penetration resistance was low at the given water content in both substrates. Further experiments are required to elucidate whether observed differences can be explained by mechanical properties beyond mechanical impedance, root or microbiome ethylene production or differences in diffusion processes within the root or in the rhizosphere. A more systematic literature review including studies comparing textures at different levels of nutrient supply is needed. Results from field studies comparing different substrates under the same environmental conditions can also help to unravel the mechanisms involved.
Theoretical considerations
Overview of the significance to root hairs in P acquisition
Root hairs are formed from either individual or a string of thin tube-like cells that grow outward perpendicular to the root epidermal surface (Brown et al. 2012b). While some studies report that root hairs assist in root water uptake (Carminati et al. 2017), root hairs are thought to develop primarily for nutrient uptake (Brown et al. 2012b). As roots develop hairs, they increase the root’s surface area, accounting for up to 70% of the total root area (Raghothama and Karthikeyan 2005). This acts to maximize P absorption potential (Brown et al. 2012b; Silberbush and Barber 1983). The geometric scale of root hairs (approximate diameter of 10 μm) enables them to exploit scarce P stocks otherwise inaccessible to the plant due to their immobility (Tisdall 1991). Furthermore, root hairs entangle and adhere to soil, creating an outer layer of aggregates known as the rhizosheath (Delhaize et al. 2009; Koebernick et al. 2017). The rhizosheath facilitates root P uptake via enhanced diffusion towards the root by reducing the tortuosity of diffusion pathways (Brown et al. 2012b; Brown et al. 2017; Pang et al. 2017). The rhizosheath forms a cluster of soil aggregates, which is abundant in microbes and mycorrhizal fungi (George et al. 2014), further interacting with the plant root and facilitate root P acquisition (Jakobsen et al. 2005). The combined effect results in root hairs accounting for over half of the total P taken up by the roots (Brown et al. 2012b; Keyes et al. 2013). Although there are many complex processes associated with enhanced P acquisition due to root hairs, the focus of this study is to assess the effect of root hairs with variation in environmental conditions at a larger scale. Therefore, this study assumes that root hairs account for half of the total P root uptake rate. Details regarding the model implementation are elaborated in the materials and methods section.
Conclusions
We developed a field scale model that considers the role that root hairs play in soil P acquisition and compares their relative impact to varying precipitation patterns and differing initial soil P quantities and distributions.
Results show that for individual growing seasons, P concentrations down the soil profile do not appear to change significantly. However, noticeable changes can be detected over several growing periods. Differences between simulations with and without root hairs suggest that, while root hairs play an important role in obtaining P, they pose negligible risk to soil nutrient mining under current agricultural procedures.
Although root hairs account for up to 50% of the total P taken up by the plant roots, increasing the P content by 70% increased the root P uptake rate by 64%. A delayed precipitation scenario reduced P uptake rate by as much as 60%. These three features all play a critical role in understanding plant roots P use efficiency.
The model results illustrate the significance of soil moisture during the growth season and suggests that irrigation strategies could be employed during the first 2 months of a given growing season to optimize the P use efficiency.
We note that all of the results from this study were based on model simulations, and we stress that more future field scale studies should be carried out to measure the impact of small scale rhizosphere features on field scale processes.
Author information
Authors and Affiliations
Department of Soil System Science, Helmholtz Centre for Environmental Research — UFZ, Theodor-Lieser-Strasse. 4, 06120, Halle/Saale, Germany
Eva Lippold, Maxime Phalempin, Steffen Schlüter & Doris Vetterlein
Soil Sciences, Martin Luther University Halle-Wittenberg, Von-Seckendorff-Platz 3, 06120, Halle/Saale, Germany
You can also search for this author in
You can also search for this author in
You can also search for this author in
You can also search for this author in
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by EL and MP. The first draft of the manuscript was written by EL, MP, SS, and DV. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Funding
Open Access funding enabled and organized by Projekt DEAL. This project was funded by DFG, German Research Foundation (project number 403801423, 403640293) as part of priority programme 2089 “Rhizosphere spatiotemporal organisation—a key to rhizosphere functions”.
")




")