

- Supplementary Material
- Impact of PGPR on Root System Architecture and Root Structure
- PGPR Effects on RSA Via Modulation of Host Hormonal Balance
- Modification of Root Cell Wall and Root Tissue Structural Properties by PGPR
- Phenotyping Platforms for Further Understanding of Root Architecture Traits
- Expression of Plant-Beneficial Functions of PGPR in the rhizosphere
- Regulation of PGPR Functions by Root Exudates
- Regulation of PGPR Functions by Microbial Signals
- Growth in Response to Soil Nutrients
- Nitrogen
- Phosphorus
- Sulfur
- Defining Root System Architecture
- Systemic Effects of PGPR on Whole Plant Physiology and Functioning
- Impact of PGPR on Plant Nutrition
- Impact of PGPR on Plant Transcriptome
- Impact of PGPR on Plant Metabolome
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/Plant_Genetics_and_Genomics/10.3389/fpls.2013.00186/abstract
Impact of PGPR on Root System Architecture and Root Structure
Most terrestrial plants develop their root system to explore soil and find nutrients to sustain growth. Root is a complex organ made of distinct regions such as the root tip, root meristem, differentiation and elongation zones, and emerging lateral roots (Scheres et al., 2002). These regions have distinct roles. For instance, root hairs are differentiated epidermal cells important for plant mineral nutrition, as inferred from gene expression studies (Lauter et al., 1996; von Wiren et al., 2000) and nutrient accumulation measurements (Ahn et al., 2004). Root functional specificity is also reflected at the level of plant-microbe interactions. In Fabaceae for example, the root tip is the most important region to initiate the rhizobial colonization process leading eventually to the formation of a root nodule (Desbrosses and Stougaard, 2011). In Poaceae, root hairs and lateral roots are preferentially colonized by PGPR, where they may express their plant beneficial properties (Pothier et al., 2007; Combes-Meynet et al., 2011). Root system architecture (RSA) integrates root system topology, the spatial distribution of primary and lateral roots, and the number and length of various types of roots. Several abiotic and biotic factors can influence RSA, including PGPR strains. PGPR modify RSA and the structure of root tissues mainly through their ability to interfere with the plant hormonal balance (Figure 1).
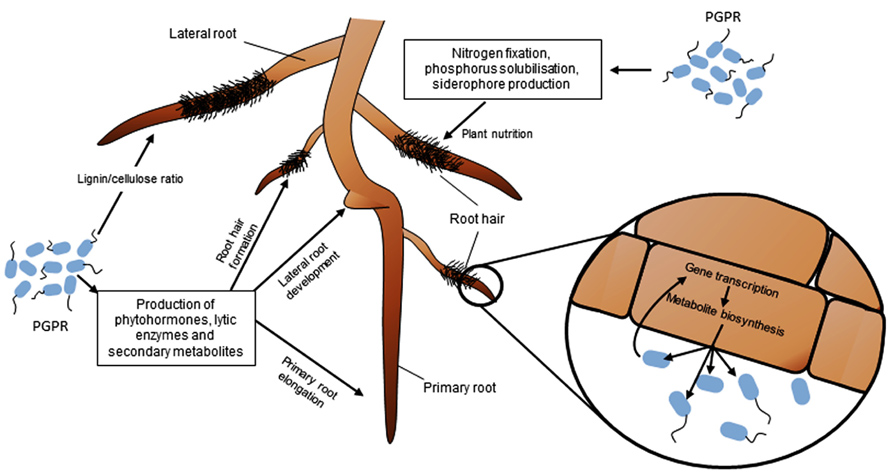
FIGURE 1. Impact of phytostimulating PGPR on RSA, nutrient acquisition and root functioning. PGPR can modulate root development and growth through the production of phytohormones, secondary metabolites and enzymes. The most commonly observed effects are a reduction of the growth rate of primary root, and an increase of the number and length of lateral roots and root hairs. PGPR also influence plant nutrition via nitrogen fixation, solubilization of phosphorus, or siderophore production, and modify root physiology by changing gene transcription and metabolite biosynthesis in plant cells.
PGPR Effects on RSA Via Modulation of Host Hormonal Balance
Changes in RSA may result from interferences of PGPR with the main hormonal pathways involved in regulating plant root development: auxin, cytokinin, ethylene, and to a lesser extend gibberellin, and abscisic acid (ABA) (Moubayidin et al., 2009; Stepanova and Alonso, 2009; Dodd et al., 2010; Overvoorde et al., 2011). The balance between auxin and cytokinin is a key regulator of plant organogenesis, and shapes root architecture (Aloni et al., 2006). The auxin to cytokinin ratio can be affected by PGPR because they are able to produce a wide range of phytohormones, including auxins and/or cytokinins, and secondary metabolites that can interfere with these hormonal pathways.
Many PGPR are able to produce phytohormones and secondary metabolites interfering with the plant auxin pathway, such as auxins, 2,4-diacetylphloroglucinol (DAPG), and nitric oxide (NO). Indole-3-acetic acid (IAA) is the best-characterized auxin produced by many plant-associated bacteria, including PGPR (Spaepen et al., 2007a). Exogenous IAA controls a wide variety of processes in plant development and plant growth: low concentrations of IAA can stimulate primary root elongation, whereas high IAA levels stimulate the formation of lateral roots, decrease primary root length and increase root hair formation (Figure 1; Dobbelaere et al., 1999; Patten and Glick, 2002; Perrig et al., 2007; Spaepen et al., 2007b; Remans et al., 2008). IAA is usually synthesized by rhizobacteria from tryptophan, which is found at different concentrations in root exudates according to plant genotype (Kamilova et al., 2006). In PGPR strains, several IAA biosynthetic pathways have been described depending on the metabolic intermediates (Spaepen et al., 2007a). The indole-3-pyruvate decarboxylase (encoded by the ipdC/ppdC bacterial gene) is a key enzyme involved in the indolepyruvic acid pathway. Effects of ipdC mutants on plant root morphology are often altered in comparison to those of wild-type strains (Brandl and Lindow, 1998; Dobbelaere et al., 1999; Patten and Glick, 2002; Suzuki et al., 2003; Malhotra and Srivastava, 2008).
Plant growth promotion by PGPR can also result from indirect stimulation of the plant auxin pathway. For example, several PGPR strains like Azospirillum brasilense have a nitrite reductase activity and consequently are able to produce NO during root colonization (Creus et al., 2005; Pothier et al., 2007; Molina-Favero et al., 2008). NO is involved in the auxin signaling pathway controlling lateral root formation (Creus et al., 2005; Lanteri et al., 2006, 2008; Molina-Favero et al., 2008). DAPG is a well-known antimicrobial compound produced by biocontrol fluorescent pseudomonads (Couillerot et al., 2009). At lower concentrations, DAPG can also be a signal molecule for plants, inducing systemic resistance (Iavicoli et al., 2003; Bakker et al., 2007), stimulating root exudation (Phillips et al., 2004), and enhancing root branching (Brazelton et al., 2008; Couillerot et al., 2011; Walker et al., 2011). DAPG can interfere with an auxin-dependent signaling pathway and thus modify RSA (Brazelton et al., 2008). Indeed, applications of exogenous DAPG, at a concentration around 10 μM, inhibited primary root growth and stimulated lateral root production in tomato seedlings. Furthermore, roots of an auxin-resistant diageotropica mutant of tomato displayed reduced DAPG sensitivity (Brazelton et al., 2008).
The growth-promotion effect of auxin or auxin-like compounds by PGPR may require functional signaling pathways in the host plant. To test that hypothesis, one could use a host plant defective at a particular step of the hormone-signaling pathway and assess whether PGPR inoculation complements or not the effect of the mutation. This strategy requires the use of model plant such as Arabidopsis, the only biological system that provides to date enough documented mutant plants (Dubrovsky et al., 1994; Alonso et al., 2003). Consistent with that, some Arabidopsis auxin-signaling mutants failed to show the typical root architecture changes in response to the beneficial rhizobacterium Phyllobacterium brassicacearum STM196 (Contesto et al., 2010). However, auxin content was not increased in roots upon inoculation with Phyllobacterium brassicacearum STM196, ruling out the potential implication of auxin of bacterial origin (Contesto et al., 2010). Nevertheless, the use of Arabidopsis DR5::GUS reporter line, whose expression is restricted to the root meristem where the auxin maximum is located (Ulmasov et al., 1997; Casimiro et al., 2001), showed a change of expression pattern in response to STM196 inoculation (Figure 2). GUS staining appeared more intense on a wider region of the root tip as well as in the vasculature, suggesting that there was a change of auxin distribution in the root in response to STM196 inoculation, even though this strain is a low auxin producer (Contesto et al., 2010). Interestingly, a similar observation was made when Arabidopsis plantlets were inoculated with the PGPR Bacillus subtilis GB03 (Zhang et al., 2007), which emits volatile organic compounds (VOCs), or with Pseudomonas fluorescens WCS417 (Zamioudis et al., 2013).
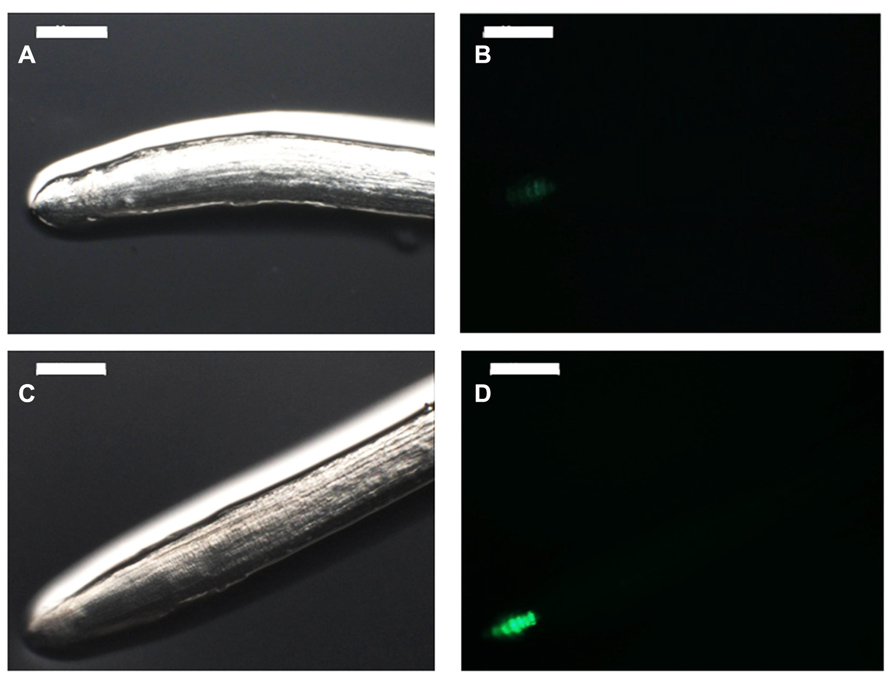
FIGURE 2. PGPR-mediated changes in RSA may relate to modifications of auxin content in roots. Six-day-old Arabidopsis plantlets expressing the GFP gene under the control of the auxin-sensitive DR5 artificial promoter were inoculated (C, D) or not (A, B) with the PGPR Phyllobacterium brassicacearum STM196. Six days later, root tips were observed under normal light (A, C) or UV light (B, D) with a microscope (Z16APO, Leica, Bensheim, Germany). Scale bars represent 200 μm. Inoculation by STM196 modified root traits such as root hair elongation and primary root growth, which coincided with an increase in GFP signal in the root tip in inoculated (D) compared with control plants (B). These observations confirm previous results with a different Arabidopsis DR5 reporter line (Contesto et al., 2010).
Cytokinin production (especially zeatin) has been documented in various PGPR like Arthrobacter giacomelloi, Azospirillum brasilense, Bradyrhizobium japonicum, Bacillus licheniformis, Pseudomonas fluorescens, and Paenibacillus polymyxa (Cacciari et al., 1989; Timmusk et al., 1999; de García Salamone et al., 2001; Perrig et al., 2007; Cassán et al., 2009; Hussain and Hasnain, 2009). Cytokinins stimulate plant cell division, control root meristem differentiation, induce proliferation of root hairs, but inhibit lateral root formation and primary root elongation (Silverman et al., 1998; Riefler et al., 2006). Inoculation of plants with bacteria producing cytokinin has been shown to stimulate shoot growth and reduce the root to shoot ratio (Arkhipova et al., 2007). Bacterial genes involved in cytokinin biosynthetic pathways have been identified in silico but their role has not yet been validated through functional analyses (Frébort et al., 2011). Consequently, the contribution of cytokinin production by PGPR to RSA modifications remains speculative.
Several reports have revealed that PGPR are able to produce ABA or gibberellic acid, or to control the level of these hormones in plants (Richardson et al., 2009; Dodd et al., 2010). The first one, ABA, is well known for its involvement in drought stress. During water stress, increase in ABA levels causes closing of stomata, thereby limiting water loss (Bauer et al., 2013). However, this hormone also plays different roles during lateral root development (De Smet et al., 2006; Dodd et al., 2010). Inoculation with Azospirillum brasilense Sp245 led to an increase of ABA content in Arabidopsis, especially when grown under osmotic stress (Cohen et al., 2008). Gibberellins promote primary root elongation and lateral root extension (Yaxley et al., 2001). Production of gibberellins has been documented in several PGPR belonging to Achromobacter xylosoxidans, Acinetobacter calcoaceticus, Azospirillum spp., Azotobacter spp., Bacillus spp., Herbaspirillum seropedicae, Gluconobacter diazotrophicus and rhizobia (Gutiérrez-Mañero et al., 2001; Bottini et al., 2004; Dodd et al., 2010). Application of gibberellic acid on maize, at a concentration similar to that produced by Azospirillum, promotes root growth; furthermore, gibberellin content increases in maize root inoculated with Azospirillum (Fulchieri et al., 1993). In addition to playing a role in plant RSA, these two hormones are involved in plant defense mechanisms. Thus, PGPR producing these hormones may modulate the hormonal balance involved in plant defense, including the jasmonate and salicylic acid pathways (for a review see Pieterse et al., 2009).
Although the production of hormones by PGPR has been well described, the genetic determinants involved in their biosynthesis remain largely unknown and bacterial mutants affected in hormone biosynthesis are mostly lacking. Consequently, the involvement of hormones of bacterial origin in the modulation of plant hormonal balance has not been fully demonstrated.
Modification of Root Cell Wall and Root Tissue Structural Properties by PGPR
Many PGPR can lead to modifications of the chemical composition and therefore structural properties of root cell walls (Figure 1; El Zemrany et al., 2007; Zhang et al., 2007). For example, the biocontrol agent Bacillus pumilus INR-7 is able to enhance lignin deposition in pearl millet epidermal tissues, and this plant defense response appears much more rapidly in PGPR-primed plants infected by the pathogen Sclerospora graminicola compared to non-primed plants (Niranjan Raj et al., 2012). The sole inoculation of INR-7 led to callose apposition. Although the precise location of these deposited polymers was not investigated, it is possible that their enhanced accumulation may participate to pathogen inhibition and disease suppression. A similar response was also triggered by Bacillus pumilus SE34 and Bacillus subtilis UMAF6639 when inoculated to respectively pea and melon roots. In both cases, it led to enhanced fungal pathogen tolerance (García-Gutiérrez et al., 2013). Inoculation of Pseudomonas fluorescens 63-28R to pea roots induced accumulation of lignin in root cells and inhibited colonization by the oomycete Pythium ultimum (Benhamou et al., 1996). The same result was observed with a Pseudomonas putida strain inoculated on bean roots (Anderson and Guerra, 1985). These cell wall modifications have been reported in the case of PGPR that protect plants against phytopathogens by activating ISR plant defense responses (Iavicoli et al., 2003; Desoignies et al., 2012; Weller et al., 2012; García-Gutiérrez et al., 2013). One of the consequences of ISR is thus the reinforcement of the cell wall through enhanced lignin synthesis and callose apposition (Kovats et al., 1991; Strömberg and Brishammar, 1993), which restricts the progression of phytopathogens through plant tissues (García-Gutiérrez et al., 2013).
Modifications of the chemical composition of root cell walls are also triggered by PGPR that directly promote plant growth (Figure 1). Through the analysis of the infrared spectral characteristics of crude cell wall preparations of maize roots, El Zemrany et al. (2007) concluded that roots inoculated with Azospirillum lipoferum CRT1 had lower lignin content than uninoculated ones. This result contrasts with those aforementioned for biocontrol agents. Nevertheless, lower lignin content may facilitate cell elongation, and therefore overall root elongation. Similarly, Azospirillum irakense produces pectate lyases that are capable of degrading the pectate content of root cell wall and might allow its progression between root cortex cells and its functioning as an endophyte (Bekri et al., 1999). Up to now, the impact on plant lignin content of PGPR that are both inducing ISR and promoting root growth has not been clarified.
Modifications of root cell wall ultrastructure are thought to result mainly from PGPR-triggered changes in plant gene expression. Indeed, Bacillus subtilis GB03 promotes Arabidopsis growth by producing VOCs that were shown to modulate the expression of 38 genes with known functions associated with cell wall structure (Ryu et al., 2003; Zhang et al., 2007). Among them, 30 were implicated in cell wall expansion or loosening. The endophytic PGPR Azospirillum irakense was also shown to stimulate the expression of polygalacturonase genes in inoculated rice roots (Sekar et al., 2000).
Chemical mediators involved in the effects of PGPR on root cell walls have received little attention. A single report has indicated that the exogenous addition of auxins enhanced the induced polygalacturonase activities observed in Azospirillum irakense inoculated rice roots (Sekar et al., 2000).
Phenotyping Platforms for Further Understanding of Root Architecture Traits
High power, high resolution GWAS and sequencing methods have far outpaced phenotyping methods necessary for the discovery of regions and underlying genes involved in plant growth and development (McNally et al., 2006; Yan et al., 2009; Huang et al., 2010; Tung et al., 2010; Zhang et al., 2010). Precise, single-trait elucidation and accurate, efficient measurement are an absolute requirement for the replicated phenotyping of large panels of individuals necessary to resolve trait–genotype associations using GWAS. Traditional methods used for root growth and architecture evaluation, such as field excavation, root bagging, plate culture, core sampling, and rhizotrons (reviewed in Shashidhar et al., 2012) are poorly suited for the large number of individuals required by GWAS due to a range of issues including low volume and sampling size, poor trait complexity resolution and measurement accuracy, and high labor, time, space, and material costs. However, these traditional approaches provide invaluable information about plant growth and yield under relevant field conditions and can be productively integrated with results from newer phenotyping platforms to provide a strong rationale for prioritizing future research.
A host of new, minimally intrusive, non-destructive, whole-root system growth systems and imaging platforms have now been developed that should revolutionize our ability to explore the genetic basic of RSA. Of these, hydroponics (Famoso et al., 2010) and gel (Fang et al., 2009; Iyer-Pascuzzi et al., 2010; Clark et al., 2011) growth systems are currently amongst those best suited for RSA trait measurement and analysis for their highly controlled and standardized rooting environments, ease in whole-root system visualization and adaptability for the imposition of environmental stresses and nutrient profiles. Both of these systems involve root growth in a non-natural, liquid or semi-solid rooting environment, however, they can require tailored adjustment for use with different plant species, and are somewhat spatially and thus developmentally limited to relatively simple root systems from small or young plants. X-ray computed tomography (Lontoc-Roy et al., 2006; Perret et al., 2007; Tracy et al., 2010), nuclear magnetic resonance (NMR; Menzel et al., 2007), laser (Braga et al., 2009), ground penetrating radar (GPR) and infrared (IR) and near-infrared (NIR) imaging systems (Tirlapur and Konig, 1999; Dokken and Davis, 2007) are advantageous in their ability to visualize plant root systems grown in soil or solid rooting media, but are currently limited by their small analysis volume and often low resolution and precision, as well as their cost, accessibility, and low-throughput.
With further advancements, NMR, GPR, and IR/NIR technologies have the greatest scale-up potential for the eventual non-destructive imaging and phenotyping of field-grown plant root systems. Although these current root growth systems and imaging technologies are still unable to accurately visualize and quantify complex, mature plant root systems grown under field conditions, they have contributed greatly to increase the precision and efficiency of 2D and 3D spatial and temporal imaging crucial for obtaining information about natural development of RSA in a solid rooting media (reviewed in Danjon and Reubens, 2007; Gregory et al., 2009). Comparative data analysis and integration, especially across controlled environment and field studies is necessary to determine whether QTLs detected by different phenotyping approaches are colocalized along the chromosomes. These regions can be targeted for further investigation to elucidate the genes and molecular mechanisms underlying the trait or phenotype(s) of interest.
The concurrent design of automated or semiautomated image capture systems and software for automated image processing, analysis, and root phenotype quantification (Armengaud et al., 2009; French et al., 2009; Famoso et al., 2010; Clark et al., 2011, 2013) are absolutely essential for simple, precise, and efficient root phenotyping with whole-root system growth platforms. These automated image capture and quantification software systems are also often easily adaptable to an array of low and high-tech growth systems, providing the potential to enhance the throughput and accuracy of root trait measurement from plants grown in a variety of growth systems. Sustained innovation inaccurate, efficient, large-scale, high-throughput root growth and analysis systems, especially those tailored toward more the complex and natural soil and field environments will continue to be essential for future studies on the association and linkage mapping of RSA traits.
Expression of Plant-Beneficial Functions of PGPR in the rhizosphere
One PGPR strain can harbor several plant-beneficial properties, which may be co-regulated or not. Within the rhizosphere, the expression of PGPR’s plant-beneficial properties is affected by both abiotic factors (like pH, oxygen, clay mineralogy, heavy metals, etc.) and biotic factors (i.e., compounds produced by plants or the rhizo-microbiome) that can lead to distinct expression patterns in space and time, possibly with different effects on host plant (Piccoli and Bottini, 1994; Pothier et al., 2008; Prigent-Combaret et al., 2008; Dutta and Podile, 2010; Almario et al., 2013b; Drogue et al., 2013). In this section, a focus is put on the regulation of the expression of PGPR plant-beneficial properties by biotic factors occurring in the rhizosphere.
Regulation of PGPR Functions by Root Exudates
Through the release of root exudates, plants can impact bacterial gene expression, especially genes encoding plant-beneficial traits. Composition of root exudates is dependent upon intra and inter-specific genetic variability (Bertin et al., 2003; Czarnota et al., 2003; Phillips et al., 2004), plant developmental stage (Lynch and Whipps, 1990; Bacilio-Jiménez et al., 2003) and soil abiotic factors (Lipton et al., 1987). One of the major studies aiming at analyzing the impact of root exudates variability on bacterial gene expression was carried out on phlA, involved in DAPG biosynthesis, in Pseudomonas protegens (formely Pseudomonas fluorescens) CHA0 (Notz et al., 2001). The expression of phlA was increased fourfold in the rhizosphere of monocots (maize and wheat) compared to the rhizosphere of dicots (bean and cucumber). The analysis of six maize cultivars also revealed that phl expression and hence biocontrol activity could be affected by plant genotype (Notz et al., 2001). Specific components of root exudates, notably sugars, were shown to affect the production of antimicrobial compounds, such as DAPG, pyoluteorin and pyrrolnitrin by fluorescent pseudomonads, with some strain-dependent effects (Duffy and Défago, 1999). Among 63 plant compounds related to defense or development, or involved in plant-microbe interactions (flavonoids, phenolic acids, phytohormones, etc.), many could modulate the expression of phlA and pltA in Pseudomonas protegens CHA0 (de Werra et al., 2011). No specific chemical structures were identified that generally induced or repressed phlA or pltA expression (de Werra et al., 2011). Umbelliferone led to the strongest inhibition of phlA; salicylate, jasmonate, and methyl jasmonate, all slightly reduced phlA expression, whereas the plant hormone IAA induced phlA expression. None of these compounds had an effect on pltA expression (de Werra et al., 2011) whereas a previous study reported repression of both DAPG and pyoluteorin biosynthesis genes by salicylate (Baehler et al., 2005).
1-Aminocyclopropane-1-carboxylic acid deamination (encoded by acdS) is another bacterial function that may be differentially expressed according to plant genotypes. Indeed, in vitro experiments demonstrated that some compounds present in root exudates tightly control acdS expression. First, ACC, the precursor of ethylene that is metabolized by AcdS, can positively regulate acdS expression (Hontzeas et al., 2004; Prigent-Combaret et al., 2008). Second, leucine, by inhibiting oligomerization of the Lrp-type regulator AcdR, prevents transcription of acdS leading to a decrease of ACC deaminase activity in Pseudomonas putida UW4 (Li and Glick, 2001) and in Azospirillum lipoferum 4B (Prigent-Combaret et al., 2008). Finally, carbon sources can also influence acdS transcription (Prigent-Combaret et al., 2008).
As presented above, bacterial IAA biosynthesis mostly depends on tryptophan-related pathways (Spaepen et al., 2007a). The main source of tryptophan for PGPR is root exudates. Measurement of tryptophan bioavailability from graminaceous roots (Avena barbata) indicated that this amino acid is abundant at the emergence of secondary roots (Jaeger et al., 1999). In the absence of exogenous tryptophan supply, bacterial IAA biosynthesis is insignificant (Ona et al., 2005; Malhotra and Srivastava, 2006). Next to being an IAA precursor, tryptophan also plays an important role in regulating positively the ipdC/ppdC gene (Ona et al., 2005). Other root-exuded amino acids like tyrosine and phenylalanine can also induce ipdC/ppdC expression (Rothballer et al., 2005). Besides amino acids, plant roots release compounds like vitamins (e.g., pyridoxine and nicotinic acid) and organic acids (e.g., phenylacetic acid and prephenic acid; Shukla et al., 2011). All these compounds increase significantly IAA production in Azospirillum brasilense Sp245 (Zakharova et al., 2000; Somers et al., 2005).
Metabolites present in root exudates can thus specifically modulate the expression of key genes involved in plant-beneficial functions. Consequently, specific physiological responses of the plant are dependent on the PGPR strain/plant cultivar combination (Drogue et al., 2012).
Regulation of PGPR Functions by Microbial Signals
Plant growth-promoting rhizobacteria exchange several types of cell-to-cell communication signals between each other and with other rhizosphere-inhabiting bacteria and fungi, i.e., quorum-sensing (QS) signals that allow bacteria to monitor their density and to coordinate gene expression only when a quorum of cells is achieved (Fuqua et al., 1994) and other bacterial signals that regulate gene expression independently of the cell density.
Quorum-sensing relies on the synthesis and perception of small diffusible molecules, such as N-acyl-homoserine lactones (AHLs). In fluorescent pseudomonads, colonization properties and biosynthesis of antimicrobial metabolites, such as phenazines, is often subjected to an AHL-based QS regulation (Pierson et al., 1994; Chin-A-Woeng et al., 2001; De Maeyer et al., 2011). Production of pyrrolnitrin in Serratia plymuthica HRO-C48, a strain isolated from the rhizosphere of oilseed rape and able to protect crops against Verticillium wilt, is also under QS regulation (Liu et al., 2007). In S. plymuthica G3, an endophytic strain, QS positively regulates antifungal activity, production of exoenzymes, but negatively regulates IAA production (Liu et al., 2011). Among the genus Azospirillum, only a few strains belonging to the lipoferum species and isolated from rice, display the ability to produce AHL signals (Vial et al., 2006). In the rice endophyte Azospirillum lipoferum B518, AHL inactivation abolishes pectinase activity, increases siderophore synthesis and reduces IAA production (in stationary phase) but no effect is observed on cellulase activity and on the phytostimulatory effect (Boyer et al., 2008). Moreover, a proteomic approach indicates that AHL-based QS regulation in Azospirillum is rather dedicated to control functions linked to rhizosphere competence and adaptation to plant roots (Boyer et al., 2008).
Interestingly, several studies have shown that bacterial communication of a specific bacterial population could be jammed by other microbes; indeed, some soil bacteria can inactivate AHL (notably members of the genus Bacillus), whereas others can intercept AHL or can act as a physical barrier preventing their diffusion (Boyer and Wisniewski-Dyé, 2009). Consequently, other members of the bacterial rhizosphere community can compromise expression of biocontrol traits in PGPR. Conversely, cross-talk between species using the same AHL signal or a structurally-related AHL can occur in natural habitats and was evidenced in the rhizosphere of wheat and tomato (Pierson et al., 1998; Steidle et al., 2001). Finally, plant compounds designated as AHL-mimics can also interfere with bacterial QS and may influence the expression of plant-beneficial functions (Teplitski et al., 2000; Vandeputte et al., 2010). Some Pseudomonas fluorescens strains unable to synthetize AHLs but possessing the cognate receptor may even recognize a plant compound to trigger expression of genes involved in biocontrol properties (Subramoni et al., 2011).
Exometabolites produced by microbial populations including pathogenic fungal strains can also affect PGPR plant-beneficial properties. For instance, fusaric acid produced by Fusarium oxysporum represses the production of DAPG in the biocontrol strain Pseudomonas protegens CHA0 (Notz et al., 2002). Next to their antifungal effect, some Pseudomonas-produced compounds can influence gene expression of biocontrol traits in pseudomonads. Indeed, in Pseudomonas protegens strains CHA0 and Pf-5, DAPG and pyoluteorin productions are influenced by positive autoregulation; moreover, DAPG and pyoluteorin mutually inhibit one another’s production (Brodhagen et al., 2004; Baehler et al., 2005). In order to determine if DAPG could act as a signal on other PGPR strains than those of the fluorescent Pseudomonas group, a differential fluorescence induction promoter-trapping approach based on flow cytometry was developed on Azospirillum. Using this approach DAPG was shown to enhance expression of a wide range of Azospirillum brasilense genes, including genes involved in phytostimulation. Four of them (i.e., ppdC, flgE, nirK, and nifX-nifB) were upregulated on roots in the presence of Pseudomonas fluorescens F113 compared with its DAPG-negative mutant (Combes-Meynet et al., 2011). Accordingly, Pseudomonas fluorescens F113 but not its DAPG-negative mutant enhanced the phytostimulatory effect of Azospirillum brasilense Sp245 on wheat. Thus, DAPG can act as a signal by which some beneficial pseudomonads may stimulate plant-beneficial activities of Azospirillum PGPR (Combes-Meynet et al., 2011). This finding is also relevant in the context of inoculation with microbial consortia, in which different types of PGPR may be combined. The number of studies investigating the efficacy of such combined inoculations is growing, with variations in the number of microorganisms and the nature of the combinations (PGPR strains only, PGPR and fungi, etc.; Cassán et al., 2009; Couillerot et al., 2012; Kumar et al., 2012; Walker et al., 2012). Field inoculation of sorghum with fluorescent Pseudomonas strains alone or in combination with arbuscular mycorrhizal fungi showed a better effect when in presence of the latter (Kumar et al., 2012). Field inoculation of maize with a consortium consisting of two PGPR (Azospirillum lipoferum CRT1 and Pseudomonas fluorescens F113) and one mycorrhizal strain (Rhizophagus irregularis/Glomus intraradices JJ291) showed an increase of root surface, root volume and number of roots, although data were not statistically significant compared to the single Rhizophagus inoculation (Walker et al., 2012). Modification of one member of this consortium (three different Azospirillum strains were tested) could lead to significant modification of maize growth (Couillerot et al., 2012). Further studies are needed to describe the synergistic effects between beneficial microorganisms at a molecular scale and to analyse the expression of plant-beneficial functions when consortia are used.
Growth in Response to Soil Nutrients
Plant root adaptive growth in response to soil macro and micronutrients depends on a wide array of variables: nutrient forms, availability, concentration, localization, and nutrient behavior in soil, as well as the nutrient status of the plant. Similar to the tropic responses above, plant root growth in response to a nutrient stimulus requires four main steps: stimulus perception, signal transduction, target gene regulation, gene product mediation of growth response.
Nitrogen
Nitrogen, the most limiting nutrient to plant growth is an interesting example of these highly plastic plant responses to nutrient availability, as it can inhibit LR outgrowth, development and elongation under high N conditions, or in soil with low inorganic nitrogen, soil patches with high inorganic nitrogen can have a local, stimulatory effect on LR elongation and branching within the high N area. Arabidopsis senses nitrate through the PR tip, with downstream components of the nitrate LR growth response pathway include high and low-affinity Arabidopsis NITRATE TRANSPORTERS1.1 and 2.1 (AtNRT1.1 and 2.1; Zhang et al., 1999; Malamy and Ryan, 2001), and nitrate-responsive TFs, including the MADS box TF ARABIDOPSIS NITRATE REGULATED1 (ANR1; Zhang and Forde, 1998).
The nitrate transporters may be either nitrate sensors or, transporters that facilitate N movement for detection via another protein. AtNRT2.1 is necessary for LR growth repression in plants with a high external carbon to nitrogen value (Malamy and Ryan, 2001; Little et al., 2005; Remans et al., 2006b), and AtNRT1.1 is a dual-affinity transporter induced by both auxin and nitrate and important for nitrate uptake under high N conditions (Liu et al., 1999; Guo et al., 2002; Munos et al., 2004). AtNRT1.1 is also an auxin influx facilitator, decreasing its auxin transport activity in response to nitrate sensing, and is proposed to repress LR development by promoting basipetal auxin transport out of LRP under low external nitrate conditions (Krouk et al., 2010). ANR1 mediates the localized N response, regulating the increased proliferation of LRs in N-dense patches, and may be a direct or indirect target of the signal perception/transduction pathway involving AtNRT1.1 (Zhang and Forde, 1998; Remans et al., 2006a). ABA may also act in the same pathway as nitrate by inhibiting LR growth under high N conditions (Signora et al., 2001; De Smet et al., 2003). SLs appear to be upregulated in plants under low N conditions (Yoneyama et al., 2007b); however, whether increased these SL levels have a definite impact on root growth has yet to be determined.
Changes in RSA may also be induced depending on the prevailing available organic form of nitrogen, such as L-glutamate or carnitine. In Arabidopsis seedlings, the sensing of L-glutamate by the PR tip inhibits cell division in the PRM and induces LR formation and outgrowth. L-glutamate may act more as a signaling molecule as opposed to a nitrogen source, as several Arabidopsis auxin-signaling mutants display varying levels of sensitivity to L-glutamate (Walch-Liu et al., 2006), and a rice glutamate receptor mutant displays a host of RSA changes, with short PR and LR, reduced cell division and RAM cell death (Li et al., 2006). Carnitine, transported in Arabidopsis by AtOCT1, has been shown to stimulate LR formation, perhaps by locally affecting the C:N ratio important in modulating LR development (Lelandais-Briere et al., 2007).
Phosphorus
Phosphorus is the second most limiting nutrient because of its high affinity to bind metals in acidic and alkaline topsoil layers, forming insoluble substrates. Phosphorus is taken up by plants as phosphate (Pi), either directly by the root system or, in arbuscular mycorrhizae hostplants, may also be transferred through the fungal symbiont – the genetic control of which will be explored in detail later in this paper.
Under high Pi conditions in Arabidopsis, PR growth is promoted, while LR growth is inhibited (Linkohr et al., 2002). Under natural conditions where Pi is limiting, plants adopt a root foraging strategy to explore topsoil layers for phosphorus. This Pi foraging strategy may be accomplished through one of several different RSA and physiological changes. In Arabidopsis and rice, growth shifts to favor an increased root:shoot ratio, with a higher initiation and outgrowth of LRs, forming a shallow, highly branched root system (Williamson et al., 2001; López-Bucio et al., 2002; Shimizu et al., 2004). Under low Pi conditions, Arabidopsis PR growth is inhibited (Williamson et al., 2001; Linkohr et al., 2002; López-Bucio et al., 2002), while root hairs increase in density and length (Bates and Lynch, 1996, 2000). In legumes, including soybean, pea, and common bean (Phaseolus vulgaris), basal root growth angle is shifted from a downward to a more horizontal direction (Bonser et al., 1996), though a recent study shows the opposite effect in Arabidopsis, with LR GSAs shifting to a steeper, downward orientation under low Pi conditions (Bai et al., 2013). Several different families of plants develop proteoid or cluster roots – highly branched bunches of LRs just below the soil surface that secrete phosphatases and organic acids which solubilize bound phosphate for uptake (Shane and Lambers, 2005; Schulze et al., 2006).
In Arabidopsis, the PR tip is the key organ involved in phosphate sensing, and the initial effect of low external Pi perception is the inhibition of PR growth by the loss of meristem activity and cell elongation (Williamson et al., 2001; Sanchez-Calderon et al., 2005). While a plant Pi-receptor has yet to be identified, studies suggest that the P5 type ATPase PHOSPHATE DEFICIENCY RESPONSE2 (PDR2), and multicopper oxidase LOW PHOSPHATE ROOT1 (LPR1) function in an endoplasmic reticulum-localized Pi-signaling pathway (Ticconi et al., 2004, 2009; Reymond et al., 2006). PHOSPHATE STARVATION RESPONSE1 (PHR1; Bari et al., 2006), an Arabidopsis MYB-like TF that binds the promoter sequences of low Pi-induced genes, and its regulator SMALL UBIQUITIN-LIKE MODIFIER1 (AtSIZ1; Miura et al., 2005), a small ubiquitin modified E3 ligase, and the downstream PHOSPHATE2 (PHO2), an E2 conjugase, and the microRNA miR-399, which regulates PHO2 expression, are all involved in Pi-deficiency-related transcriptional changes (Bates and Lynch, 2000; Bari et al., 2006). The Arabidopsis WRKY75 TF is also induced during Pi-deprivation and may modulate both phosphate and non-phosphate induced LR development and control the transcription of genes such as high-affinity Pi transporters important for Pi uptake (Devaiah and Raghothama, 2007; Devaiah et al., 2007). The Pi-induced tobacco bZIP TF PHOSPHATE INDUCED2 (PHI2; Sano and Nagata, 2002) and rice bHLH TF PI STARVATION-INDUCED TRANSCRIPTION FACTOR1 (OsPTF1; Yi et al., 2005) may also have a role in modulating low Pi-induced changes in RSA.
Increased auxin sensitivity, decreased CK sensitivity, and changes in auxin transport and localization appear to be at least partially responsible for Pi stress-induced LR development. A shift in auxin overaccumulation from the PR apex to the LRP, or an increased sensitivity of LRP to auxin have been suggested as proposed mechanisms for increases in LRP emergence and LR density (Franco-Zorrilla et al., 2002; López-Bucio et al., 2005; Nacry et al., 2005). TIR1 auxin receptor-dependent degradation of TF-repressing AUX/IAA proteins is essential for LR development in Pi-stressed seedlings (Perez-Torres et al., 2008). The effect of auxin under low Pi conditions is also regulated by CK signaling, which represses auxin-induced gene transcription Pi-starved Arabidopsis plants display a decreased response to CK, partly due to the reduced expression of the CR receptor CRE1 (Franco-Zorrilla et al., 2002). Ethylene perception is likely also necessary for increased root hair development and LR elongation and decreased PR elongation under low Pi conditions (Schmidt and Schikora, 2001; López-Bucio et al., 2002; Ma et al., 2003) and has additionally been shown to affect Pi stress-induced changes in basal root growth angle in bean (Lynch and Brown, 2001). Similar to CK, GA acts as a negative repressor of Pi-induced root architecture changes under low Pi conditions; Pi-deficient plants accumulate DELLA proteins, which repress GA-induced root growth suppression and thus allow for auxin-mediated LR initiation and elongation (Jiang et al., 2007). SL production is induced by low Pi in many species including tomato, Arabidopsis, pea, and rice (López-Ráez and Bouwmeester, 2008; Umehara et al., 2010; Kohlen et al., 2011; Foo et al., 2012; Mayzlish-Gati et al., 2012). Some studies suggest that increased production and exudation of SLs under soil Pi or N deficiency is dependent on whether the plant (1) is an arbuscular mycorrhizal fungi (AMF)-compatible host, and (2) whether it is dependent on the arbuscular mycorrhizal symbiosis (AMS) for Pi and N uptake (Yoneyama et al., 2007a,b, 2008; Umehara et al., 2010); however, what effect, if any, this increased SL exudation has on root growth is unclear. Exogenously applied GR24 appears to increase LR formation under low Pi or decrease LR formation under sufficient Pi though the F-box protein MORE AXILLARY GROWTH2 (MAX2), a putative component of the SL-signaling pathway (Kapulnik et al., 2011; Ruyter-Spira et al., 2011).
Sulfur
Sulfur, taken up by plant roots as sulfate, is another limiting plant macronutrient, and is essential for the synthesis of methionine and cysteine. Sulfur deficiency can have significant effects on RSA; sulfate limited Arabidopsis and maize plants increase their LR production, developing an extensive, highly branched root system, often at the expense of shoot growth (Kutz et al., 2002; Bouranis et al., 2008). Another conflicting Arabidopsis study found a decrease in LRP and emerged LR under low-sulfate growth conditions (Dan et al., 2007). To rectify these two opposing developmental outcomes, a two-state model was proposed wherein short-term sulfur limitation let to increased LR growth for sulfate foraging, but longer-term sulfate deficiency led to overall decreased growth and photosynthesis, ending in premature senescence (Hoefgen and Nikiforova, 2008; Lewandowska and Sirko, 2008).
While the genes involved in internal and external sulfate sensing and transcriptional regulation have not yet been cloned and characterized, several components of root sulfate import and signal transduction have been identified. Of the five major classes of sulfate transporters identified in Arabidopsis and rice (Takahashi et al., 1999; Buchner et al., 2004; reviewed in Takahashi, 2010), the group 1 high-affinity transporters are essential for root sulfate uptake. Arabidopsis SULFATE TRANSPORTER1;2 (SULTR1;2) is expressed under both sulfate-sufficient and low-sulfate conditions and transcriptionally regulated by the ETHYLENE-INSENSITIVE3-LIKE3 TF SLIM1, whereas the SULTR1;1 gene induced only under sulfate stress (Takahashi et al., 2000; Yoshimoto et al., 2002; Maruyama-Nakashita et al., 2004) and upregulated by O-acetylserine (thiol) lyase (OAS), a rate-limiting enzyme involved in sulfate assimilation into cysteine (Leustek et al., 2000; Saito, 2000).
Auxin may play a central role in LR production under sulfate stress. In Arabidopsis, sulfate deficiency activates the transcription of NITRILASE3 (NIT3), which converts indole-3-acetonitrile to the auxin IAA (Kutz et al., 2002). However, while NIT3 activity is especially upregulated in LRP under sulfate limitation, increased concentrations of auxin have not been proven (Kutz et al., 2002; Lewandowska and Sirko, 2008). Studies of sulfur-limitation regulated auxin signaling genes such as BIG, named for the huge 560 kD calossin-like protein it encodes, required for the polar transport of auxin (Gil et al., 2001), as well as the auxin TF genes IAA13, IAA28, and ARF-2, indicate that auxin is likely involved in the indirect regulation of sulfur homeostasis and short to long-term sulfur deficiency responses (Hirai et al., 2003; Maruyama-Nakashita et al., 2003; Nikiforova et al., 2003; Hoefgen and Nikiforova, 2008; Lewandowska and Sirko, 2008). JA may also play a role in sulfur regulation, as demonstrated by research in Arabidopsis finding low sulfur JA biosynthesis genes upregulated under low sulfur in (Maruyama-Nakashita et al., 2003), exogenous application of JA promoted sulfur assimilation and there is also evidence to suggest that CKs and sucrose may affect sulfur responsive gene transcription (Ohkama et al., 2002).
Defining Root System Architecture
Root system architecture is a complex trait and refers to the spatial configuration of the root system in terms of the precise geometric arrangement of all root axes as laid down in the rooting medium. Root architecture is comprised of a whole system set of descriptors, and as such is senior to and distinct from, though naturally dependent on, the secondary fields of root anatomy, morphology, topology, and distribution; however, individual root architecture components may draw on or overlap with these fields. To clarify, root anatomy refers to the internal cellular structure and arrangement of a root; root morphology, the surface features, including diameter, root hair and cap characteristics, and contorsion; root topology, the hierarchical description of the connection of root axes to one another; and root distribution, the presence and distribution of roots in a positional gradient or grid along a horizontal and/or vertical axis.
As proposed by Fitter (1991), there are five main components of root architecture, each of which may be comprised of several specific traits or parameters. These components are: (1) branch magnitude – the number of interior links (internode segments between two branching points or nodes) or exterior links (internode segments between a branching point and an endpoint, i.e., root apical meristem (RAM); (2) topology, the pattern of branch distribution, which is usually herringbone (alternate lateral branching off a parent root), dichotomous (opposite, bifurcating branches), or radial (whorls of branches around a parent root (Hochholdinger, 2009; Lynch and Brown, 2012); (3) link/internode lengths, the distance between branch points among different root orders of an individual root, which may be averaged across a system; (4) root angles, specifically the azimuth (radial angle) of a lateral root’s (LR) emergence around the circumference of a parent root, the branching angle or departure rate of a LR from a parent root, and the spreading angle of the entire system; and (5) link radius, the diameter of any given root (Fitter, 1991).
Systemic Effects of PGPR on Whole Plant Physiology and Functioning
In addition to their effects on root tissues, PGPR can modify the physiology and functioning of plant tissues located at a substantial distance from the colonized sites, such as shoots. Two types of mechanisms are involved. On the one hand, some PGPR can enhance nutrient availability/uptake for plant roots. Stimulation of plant nutrition will lead to modifications in primary metabolism and consequently will contribute to enhance growth. On the other hand, certain PGPR trigger specific systemic responses, mostly by unknown signaling mechanisms. High-throughput analyses of plant transcriptomic and metabolomic responses have evidenced the effects of PGPR on plant gene expression and metabolite accumulation, respectively. These results highlight the extensive effect of PGPR on whole plant physiology and functioning (Figure 1), and provide clues to understand the systemic effect of PGPR.
Impact of PGPR on Plant Nutrition
The impact of PGPR on plant nutrition may result from effects on plant nutrient uptake and/or on plant growth rate (Mantelin and Touraine, 2004). It is indeed commonly hypothesized that nutrient uptake is increased as a consequence of increased root surface area triggered by PGPR. However, root ion transporters are under the control of regulatory processes that adjust their activity to the plant nutritional demand (Imsande and Touraine, 1994; Lappartient and Touraine, 1996; Lappartient et al., 1999; Nazoa et al., 2003), so that regulations of root development and ion transporter activities are antagonistically coordinated to maintain steady nutrient acquisition rate (Touraine, 2004). Hence, PGPR must interfere with pathways coordinating plant development and plant nutrition to elicit both increased nutrient acquisition rate and plant growth promotion (Figure 1).
Plant growth-promoting rhizobacteria can directly increase nutrient supply in the rhizosphere and/or stimulate ion transport systems in root. With regards to increased nutrient supply, two main types of bacterial activities can be considered. Firstly, phosphate solubilization is one key effect of PGPR on plant nutrition. Soils generally contain a large amount of phosphorus, which accumulates in the wake of regular fertilizer applications, but only a small proportion of the latter is available for plants. Plants are able to absorb on their own mono and dibasic phosphate; organic or insoluble forms of phosphate need to be mineralized or solubilized by microorganisms, respectively (Richardson et al., 2009; Ramaekers et al., 2010). Many PGPR – such as Pseudomonas, Bacillus, Rhizobium – are able to dissolve insoluble forms of phosphate (for a review see Richardson et al., 2009). Two main processes exist: acidification of the external medium through the release of low molecular weight organic acids (such as gluconic acid) that chelate the cations bound to phosphate (Miller et al., 2009), and production of phosphatases/phytases that hydrolyse organic forms of phosphate compounds. Secondly, many associated bacteria can fix N2 so that they could provide nitrogen to the plant. Evidence in favor of the participation of PGPR to the plant N budget has been reported for several plants, especially sugar cane (Boddey et al., 2003). However, the impact of N2-fixation by PGPR is still debated and is rarely credited for the stimulation of plant growth (for review see Dobbelaere et al., 2003). In addition, non-fixing rhizobacteria can promote plant growth, showing that N provision is not obligatory for plant growth promotion. For instance, Phyllobacterium brassicacearum STM196 is unlikely to fix N2 while it promotes the growth of canola and Arabidopsis (Bertrand et al., 2000, 2001; Mantelin et al., 2006).
In Arabidopsis, NO3− uptake measurement in response to PGPR, over time, can lead to contradictory results: NO3− influx was increased in seedlings, upon 24 h-inoculation with Phyllobacterium brassicacearum STM196, while it was reduced 7 days later (Mantelin et al., 2006). However, it is hard to draw a firm conclusion as the net NO3− uptake rate remained unknown since ion efflux was not measured in these experiments. Except for the NRT2.5 and NRT2.6 genes, the accumulation of transcripts of nitrate and ammonium transporters was very slightly or not significantly changed upon Phyllobacterium brassicacearum STM196 inoculation (Mantelin et al., 2006). Interestingly, these two genes have recently been shown to be required in Arabidopsis growth promotion by this PGPR (Kechid et al., 2013). Since these two genes code for two plasma membrane-localized NO3− transporters (Kotur et al., 2012), this discovery raises the question of the interactions between N nutrition and plant development in PGPR-inoculated plants. The NRT2.5 and NRT2.6 genes are predominantly expressed in shoots (Mantelin et al., 2006). Their role in Phyllobacterium brassicacearum STM196 plant growth promotion and/or root architecture modification are not linked to changes in NO3− uptake rate or NO3− distribution between roots and shoots (Kechid et al., 2013), suggesting an involvement in N-signaling rather than a direct role in N-metabolism.
Evidence in favor of a regulation of ion transporters at a transcriptional level by PGPR has been obtained in studies with Bacillus subtilis GB03. This strain induces concomitant down- and up-regulation of HKT1 expression in roots and shoots of Arabidopsis seedlings, respectively (Zhang et al., 2008). In the shoots, HKT1 functions in phloem tissues to retrieve Na+ from the xylem (Berthomieu et al., 2003) and in the roots it is involved in Na+ uptake (Rus et al., 2001). The differential regulation of HKT1 expression in roots and shoots resulted in reduced accumulation of Na+ and increased accumulation of K+ in both organs of GB03-inoculated seedlings under salt-stress conditions (Zhang et al., 2008). Consistent with the effect of GB03 on HKT1, GB03 failed to rescue salt-stressed hkt1 mutant seedlings from elevated Na+ accumulation.
Volatile organic compounds emitted by GB03 also activate the plant’s iron acquisition machinery leading to increased iron assimilation (Zhang et al., 2009). Firstly, this PGPR leads to acidification of the rhizosphere, both directly due to chemical effects of some unidentified VOCs and indirectly through increased root proton efflux. Secondly, GB03 up-regulates the expression levels of FRO2 and IRT1 genes, coding respectively for a Fe3+ chelate reductase and a Fe2+ transporter. As a result, GB03-exposed Arabidopsis has enhanced ferric chelate reductase activity and increased iron content. Finally, it has been shown that this PGPR induces the expression of the FIT1 transcription factor that regulates positively FRO2 and IRT1 expressions (Zhang et al., 2009). The fact that GB03 fails to increase root ferric reductase activity and plant iron content in Arabidopsis fit1 mutants shows that PGPR can modify indirectly ion uptake by interfering with plant regulatory processes that control ion transporter expressions and/or activities (Zhang et al., 2009).
Impact of PGPR on Plant Transcriptome
As for endophytes, differential colonization of rice roots was observed with an Azoarcus PGPR. In a less compatible interaction, a slight defense response occurred and was accompanied by the induction of pathogenesis-related proteins and proteins sharing domains with receptor-like kinases induced by pathogens; those proteins were also induced by a jasmonate treatment (Miché et al., 2006). Inoculation of rice roots with the endophytic PGPR Herbaspirillum seropedicae triggered the expression of genes responsive to auxin and ethylene and the repression of the defense-related proteins PBZ1 and thionins (Brusamarello-Santos et al., 2012). These studies suggest that endophytes modulate plant defense responses during colonization.
Plants treated with biocontrol PGPR, usually belonging to the Pseudomonas genus, are more resistant to subsequent infections by bacterial or fungal pathogens. In Arabidopsis, this rhizobacteria-mediated ISR requires sensitivity to jasmonate and ethylene, and the regulators MYC2 (Pieterse et al., 1996, 2000; Pozo et al., 2008), NPR1 (Pieterse et al., 1998), and MYB72 (Van der Ent et al., 2008) played a central role in this signaling. One of the earliest transcriptomic study performed with Pseudomonas fluorescens WCS417r indicated that bacteria elicited a substantial change in the expression of 97 genes in roots whereas none of the approximately 8,000 genes tested showed a consistent change in expression in the leaves (Verhagen et al., 2004). Subsequent studies on Arabidopsis reported an increase of defense-related transcripts, including PR-related proteins, in shoots of bacterized plants compared to untreated shoots (Cartieaux et al., 2003; Wang et al., 2005; van de Mortel et al., 2012). Interestingly, the ISR induced by Pseudomonas fluorescens SS101 was recently reported to be dependent on salicylic acid signaling and not on jasmonic acid and ethylene signaling (van de Mortel et al., 2012); moreover, a prominent role of camalexin and glucosinolates in the ISR was proposed. In wheat, bacterization with Pseudomonas fluorescens Q8r1-96 also triggered the accumulation of defense-related transcripts (Okubara et al., 2010; Maketon et al., 2012) and neither DAPG nor the type three secretion system were key single factors in the expression of these genes (Maketon et al., 2012). The establishment of beneficial associations requires mutual recognition and substantial coordination of plant and microbial responses and consequently beneficial microbes modulate plant immunity.
Impact of PGPR on Plant Metabolome
Several studies have addressed the metabolomic changes triggered by PGPR inoculation, by analyzing metabolite contents of root exudates, root tissues and shoots under normal or stressful conditions (Figure 1). Some studies have shown that PGPR can elicit changes in the activities of root enzymes involved in the production of metabolites, especially flavonoids, leading to changes in the pattern of root exudation (Lavania et al., 2006; Shaw et al., 2006). Some Azospirillum PGPR stimulated by up to one-third the level of carbon compounds exuded from roots (Heulin et al., 1987). Moreover, compounds of microbial origin, such as phenazines and DAPG, could enhance total net efflux of amino acids in plant species (Phillips et al., 2004). On soybean roots, the PGPR Chryseobacterium balustinum Aur9 influences flavonoids exudation (Dardanelli et al., 2010). PGPR strains from Chryseobacterium (Dardanelli et al., 2010) or Azospirillum (Burdman et al., 1996) may influence flavonoid exudation by Fabacea roots. This property can be important for the design of mixed inoculants that will include a PGPR strain promoting flavonoid exudation together with rhizobia that will respond to plant flavonoids (Burdman et al., 1996).
Accumulation of secondary compounds was also modified in several plants inoculated with consortia containing arbuscular mycorrhizal fungus and PGPR. Blumenin accumulation triggered by Rhizophagus irregularis (formerly Glomus intraradices) in barley and wheat roots was increased when a rhizosphere bacterium was applied with the fungus (Fester et al., 1999). Leaf secondary metabolites (total phenols and ortho dihydroxy phenols), as well as leaf mineral content (phosphorus, potassium, zinc, copper, and iron) were maximal when Begonia malabarica or Solanum viarum were inoculated with consortia containing two fungi and a Bacillus coagulans strain (Selvaraj et al., 2008; Hemashenpagam and Selvaraj, 2011). Field-inoculation of maize with selected strains of Pseudomonas, Azospirillum or Rhizophagus/Glomus, or with these strains combined two by two or all three together, led to qualitative and quantitative modifications of root secondary metabolites, particularly benzoxazinoids and diethylphthalate (Walker et al., 2012). These modifications depended on fertilization level and on the type of microorganisms inoculated. The three selected strains gave distinct results when used alone, but unexpectedly all microbial consortia gave somewhat similar metabolic responses.
Plant growth-promoting rhizobacteria can help plants to withstand saline stress, a feature that may be linked to accumulation of specific metabolites. A higher level of proline content was reported in inoculated Bacopa monnieri (Bharti et al., 2013), as well as higher accumulation of glycine betaine-like quaternary compounds in rice inoculated with Pseudomonas pseudoalcaligenes (Jha et al., 2011). Similarly, Arabidopsis inoculation with the VOC-emitting strain Bacillus subtilis GB03 induced strong plant accumulation of glycine betaine and its precursor choline, and GB03-induced drought tolerance was lost in the xipotl mutant of Arabidopsis with reduced choline production (Zhang et al., 2010). Alleviation of cold stress was demonstrated for Burkholderia phytofirmans PsJN on grapevine; this endophytic strain promotes plant post-chilling recovery by improving acclimation to cold (Ait Barka et al., 2006). This is accompanied by accumulation of stress-related metabolites such as proline, malondialdehyde and aldehydes (known as lipid peroxidation markers), hydrogen peroxide, and by higher expression of defense- and cold-related genes (Theocharis et al., 2012). Bacterization resulted in a 1.2-fold increase in starch content and in a two-fold increase in total soluble sugars, with sugars known to be involved in low-temperature tolerance (glucose, sucrose, and raffinose) displaying higher concentrations in treated plantlets (Fernandez et al., 2012). Independently of temperature, inoculation also enhanced phenolic content (Ait Barka et al., 2006).
")




")